

1 Introduction
The genus Ananteris Thorell, 1891 is a group of scorpions with a Gondwanan distribution [1] currently known from 62 species mostly from Venezuela and Brazil [2–7]. In Colombia four species were known until recently: Ananteris columbiana Lourenço, 1991, Ananteris ehrlichi Lourenço, 1994, Ananteris gorgonae Lourenço and Flórez, 1989 and Ananteris leilae Lourenço, 1999 [4,8]. Recent contributions increased the richness of this genus in the country to eight species, with the descriptions of Ananteris myriamae Botero-Trujillo, 2007, Ananteris tolimana Teruel and García, 2007 [5,7], and two new species described by Botero-Trujillo [6].
In the present paper two additional new species are described from El Tuparro Natural National Park, Llanos ecoregion of eastern Colombia (Fig. 1), bringing to 64 the known species of Ananteris, ten of which are present in Colombia. Both new species were collected during a research project by the Instituto de Investigación de Recursos Biológicos Alexander von Humboldt on the biological characterization of the northeastern section of the Natural Park. Such study was aimed at characterizing landscapes, plants and certain animal groups among which arachnids were not included, and resulted on a recently published book [9]. Taking into account that the type locality of both new species is near the border with Venezuela, it is plausible that these may occur too in the neighboring country at the northwestern region of the State of Amazonas, a state where Ananteris is unrecorded (see [2]: 216; [10]: 60, 62; [11]).

Known distribution of Colombian Ananteris. The expanded area in gray corresponds to El Tuparro Natural National Park.
The recent increase in the known diversity of this genus, resulting from various contributions where 32 species were described [2–7,12], and the two new species described herein, suggest that the inventory work of the genus Ananteris is far from completed. Therefore, it is important to note the need of carrying on further collecting and exploration in the poorly known regions of tropical South America, and using appropriate collection techniques such as pitfall, Winkler, and nocturnal searches with ultraviolet light, in order to identify populations and species of Ananteris that may have remained unrecorded and thus better ascertain the distribution and diversity of this genus.
2 Methods
Photographs were taken using an Olympus D-590 ZOOM digital camera. Illustrations were prepared with the aid of a camera lucida mounted onto a Zeiss Stemi SV 6 stereoscope. Measurements (L = length, W = width, D = depth) were obtained following the methodology of Sissom et al. [13], using a stereoscope equipped with grid ocular micrometer. The distribution map was produced with the program ArcView GIS version 3.1. [Environmental Systems Research Institute (ESRI), Redlands, California.] Specimens are preserved in 70% ethanol.
General carinal terminology follows Vachon [14], except for metasomal carinae after Prendini [15,16], and tergal carinae that are here distinguished as follows: axial, dorsolateral and lateral carinae. Vachon's [14] term ventrointernal to denote the carina that follows the dorsointernal on pedipalp femur is here replaced for internal median, since in the specimens studied herein there is an additional and more ventral carina to which the term ventrointernal is more suitable. Trichobothrial terminology after Vachon [17,18]. The diagnoses of the new species were prepared after comparison with specimens of other Colombian species and judging from the literature available.
The specimens examined for this study are lodged in the following museums: Instituto de Investigación de Recursos Biológicos Alexander von Humboldt, Villa de Leyva, Colombia (IAvH); Museo Javeriano de Historia Natural “Lorenzo Uribe S. J.”, Pontificia Universidad Javeriana, Bogotá, Colombia (MPUJ); Instituto de Ciencias Naturales, Museo de Historia Natural, Universidad Nacional de Colombia, Bogotá, Colombia (ICN-MHN).
3 Taxonomy
Family Buthidae Koch, 1837
Genus Ananteris Thorell, 1891
Ananteris faguasi sp. nov.

Ananteris faguasi sp. nov. male holotype (IAvH-E 100786). 2. Dorsal view. 3. Ventral view. Scale bars equal 5 mm. (For interpretation of the references to colour in this figure, the reader is referred to the web version of this article.)
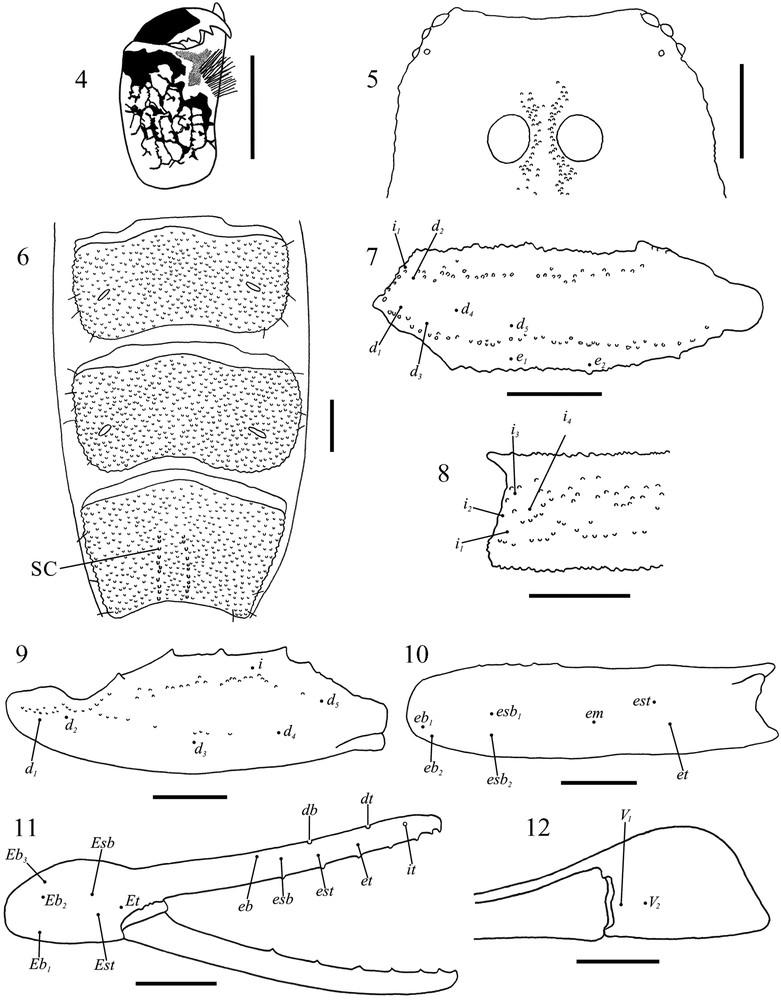
Ananteris faguasi sp. nov., male holotype (IAvH-E 100786). 4. Dorsal view of left chelicera. 5. Anterior margin of the carapace (only granulation of the median ocular carinae is included). 6. Sternites V–VII. 7–12. Distribution of the trichobothria (illustrated from the right pedipalp). 7. Femur, dorsoexternal view. 8. Femur, internal view. 9. Patella, dorsal view. 10. Patella, external view. 11. Chela, external view. 12. Chela, ventral view. SC = Submedian Carina. Scale bars equal 0.5 mm.
Meristic data for the type specimens of Ananteris faguasi sp. nov. and Ananteris volschenki sp. nov. Measurements are given in millimeters
| Ananteris faguasi sp. nov. | Ananteris volschenki sp. nov. | ||||
| Male holotype (IAvH-E 100786) | Male paratype (IAvH-E 100787) | Male holotype (IAvH-E 100788) | Female paratype (IAvH-E 100789) | ||
| Total body L (including telson) | 19.48 | 18.75 | 19.06 | 20.66 | |
| Carapace | Length | 2.38 | 2.30 | 2.38 | 2.64 |
| Anterior W | 1.04 | 0.96 | 0.93 | 1.36 | |
| Posterior W | 2.13 | 2.04 | 2.10 | 2.73 | |
| Ocular diameter | 0.21 | 0.78 | 0.21 | 0.21 | |
| Interocular distance | 0.16 | 0.16 | 0.14 | 0.19 | |
| Mesosoma | Total L | 4.96 | 4.62 | 3.84 | 4.18 |
| Metasoma | Total L (including telson) | 12.14 | 11.83 | 12.82 | 13.84 |
| Segment I L | 1.23 | 1.22 | 1.50 | 1.28 | |
| Segment I W | 1.36 | 1.20 | 1.12 | 1.78 | |
| Segment I D | 1.12 | 1.04 | 1.04 | 1.58 | |
| Segment II L | 1.50 | 1.44 | 1.63 | 1.86 | |
| Segment II W | 1.20 | 1.17 | 1.09 | 1.78 | |
| Segment II D | 1.20 | 1.06 | 1.04 | 1.50 | |
| Segment III L | 1.60 | 1.54 | 1.63 | 2.00 | |
| Segment III W | 1.23 | 1.14 | 1.09 | 1.78 | |
| Segment III D | 1.20 | 1.09 | 1.12 | 1.60 | |
| Segment IV L | 2.35 | 2.38 | 2.33 | 2.40 | |
| Segment IV W | 1.20 | 1.09 | 1.09 | 1.70 | |
| Segment IV D | 1.12 | 1.07 | 1.09 | 1.65 | |
| Segment V L | 2.91 | 2.85 | 3.09 | 3.36 | |
| Segment V W | 1.12 | 1.09 | 1.20 | 1.64 | |
| Segment V D | 1.12 | 1.10 | 1.12 | 1.68 | |
| Telson L | 2.55 | 2.40 | 2.64 | 2.94 | |
| Vesicle W | 0.64 | 0.62 | 0.56 | 0.85 | |
| Vesicle D | 0.72 | 0.67 | 0.56 | 0.78 | |
| Pedipalps | Total L | 6.98 | 7.06 | 7.53 | 9.37 |
| Femur L | 2.00 | 1.88 | 2.13 | 2.49 | |
| Femur W | 0.51 | 0.58 | 0.53 | 0.72 | |
| Patella L | 2.28 | 2.38 | 2.55 | 2.88 | |
| Patella W | 0.64 | 0.67 | 0.70 | 1.04 | |
| Chela L | 2.70 | 2.80 | 2.85 | 4.00 | |
| Chela W | 0.48 | 0.48 | 0.43 | 0.67 | |
| Chela D | 0.48 | 0.48 | 0.46 | 0.64 | |
| Movable finger L | 2.13 | 2.13 | 2.45 | 3.15 | |
| Palm L | 0.69 | 0.64 | 0.64 | 0.80 | |
| Pectines | Teeth count | 19:19 | 17:17 | 15:15 | 15:16 |
Type material.Holotype: COLOMBIA: Vichada department: adult male from Cumaribo, Santa Rita, Parque Nacional Natural El Tuparro, N, W, 135 m (260 m sensu Villareal-Leal [19]), Bosque de Cerro, trap for flight interception, 18 February 2004, I. Quintero and E. González (IAvH-E 100786).
Paratype: COLOMBIA: Vichada department: one adult male from Cumaribo, Santa Rita, Parque Nacional Natural El Tuparro, N, W, 135 m (260 m sensu Villareal-Leal [19]), Bosque de Cerro, trap for dung beetles, 18–20 February 2004, I. Quintero and E. González (IAvH-E 100787).
Etymology. Patronym dedicated to Giovanny Fagua, director of the Laboratorio de Entomología of the Pontificia Universidad Javeriana (Bogotá), in recognition of his academic labor and acknowledgment of his teachings in entomology.
Diagnosis (based on male only).Ananteris faguasi sp. nov. differs from all other species of the genus by the following unique combination of features: and trichobothria are aligned axially (Fig. 12); est and et trichobothria are located between db and dt on pedipalp fixed finger, and esb is basal to db (Fig. 11); the anterior margin of the carapace is moderately, evenly concave (not bi-concave) (Fig. 5); the median ocular carinae are granulose (Fig. 5); the metasomal carinal formula is 10:10:8:8:5 with all carinae complete; the dorsal surface of chelicerae exhibits a complete and very reticulated pattern (Fig. 4); sternite VII presents incomplete submedian carinae (Fig. 6); pectines have 17–19 teeth; and sternites IV–VII are densely granulose (Fig. 6).
Description of the male holotype (IAvH-E 100786).
Coloration. General coloration brownish with variegated pigmentation over almost the entire body (except ventrally) and appendages. Carapace predominantly dark-brown with some yellowish spots and bands; anterior and posterior margins brown; arising posterior to each lateral ocular tubercle there is a thin and almost straight yellowish line directed toward the midline, but ending behind and beside the median ocular tubercle; median ocular tubercle black. Chelicerae with coxa predominantly yellow; hand with a complete reticular pattern on dorsal surface; movable finger with a large dorsal brown area and reddish teeth; fixed finger yellow, with inconspicuous brown spots basally and reddish teeth (Fig. 4). Coxosternal region, genital operculum, pectinal basal piece, pectines and sternites III–V completely yellow, without marks; sternite VI with inconspicuous brown spots posteriorly and laterally; sternite VII with light-brown areas between the midline and the lateral margins; sternite III with a triangle-like posterior median hyaline area. Tergites predominantly brown; I–VI with a yellow longitudinal area at each side of the midline, only present anteriorly, behind which a yellow arrow-like design is present; each side of tergites I–VI with two transverse yellow lines converging near the longitudinal lines, arrow-like; tergite VII with variegated pigmentation laterally, predominantly yellow dorsally but with a brown longitudinal area on the place of the axial carina; lateral margins of tergites I–VII yellow. Metasoma predominantly brown; dorsal intercarinal spaces of segments I–IV predominantly yellow, brownish medially; segments I–IV with brown mottling throughout, except ventrally on segments I–III where predominantly yellow; segment V dark-red, lighter anteriorly. Telson reddish, lighter than segment V; aculeus slightly yellowish basally; subaculear tubercle yellow. Pedipalps predominantly brown; coxa and trochanter with variegated pigmentation; femur brownish in dorsal and internal surfaces, with a longitudinal band externally and predominantly yellow ventrally, insertion of all the trichobothria surrounded by a rounded yellow area; patella with variegated pigmentation in dorsal, external and ventral surfaces, with light-brown areas, insertion of all the trichobothria yellow; hand completely yellow in all surfaces; fixed and movable fingers brownish-yellow. Legs with variegated pigmentation, brown spots on all segments except for the telotarsus that is completely yellow.
Morphology. Measurements in Table 1. Carapace: densely covered with large rounded granules; lateral margins not parallel; anterior margin moderately, evenly concave (not bi-concave) (Fig. 5); median ocular carinae evident, others inconspicuous; a median depression anterior to the ocular tubercle and a posterior median longitudinal furrow are evident; median ocular tubercle low, located on the posterior portion of the anterior third of the carapace, with abundant granulation on the median ocular carinae; lateral ocular tubercles each with four ocelli, one of which is very reduced and unaligned with the remaining three (Fig. 5). Chelicerae: with abundant setae on the internal and ventral surfaces; cheliceral dentition characteristic of the family Buthidae [20]. Movable finger externally with two small basal teeth, one median pronounced, one subdistal slightly shorter than the median, and one distal tooth. Internally with two pronounced teeth, one basal and one median, and one distal tooth. Fixed finger externally with one basal and one median tooth mounted onto a bicuspid, one subdistal, and one distal tooth. Internally with only one small tooth located slightly basal in respect to the external subdistal. Coxosternal region: with few sparse setae; sternum subtriangular, with a deep median depression; all the components of this region almost completely smooth, except for coxae II–IV that possess scarce weak granules anteriorly; coxapophyses I–II with dense pilosity anteriorly. Genital operculum and pectines: genital operculum divided longitudinally; pectinal basal piece as long as wide; pectines long, surpassing the lateral margins of sternite III (Fig. 3); segments count on the pectines: basal lamellae 3:3, middle lamellae 8:9, teeth 19:19, fulcra absent. Sternites: sternite III almost completely smooth, except for some weak granules on the posterolateral margins; IV with abundant granulation; V–VII densely granulose throughout (Fig. 6); VII with incomplete parallel vestiges of submedian carinae, difficult to distinguish from the remaining granules, and lateral carinae completely absent; all sternites with few setae; spiracles oval elongate (Fig. 6). Tergites: with similar granulation to that of the carapace; axial carina only evident in the posterior half of tergites I–VI; dorsolateral carinae absent; tergite VII tetracarinate (paired dorsolateral and lateral carinae, incomplete), a median elevation is present on the position of the axial carina. Metasoma: with few setae; all carinae complete and serrulose; segments I–II with ten carinae (paired ventrosubmedian, ventrolateral, median lateral, dorsolateral and dorsosubmedian carinae); segments III–IV with eight (median lateral carinae absent); segment V with five (ventromedian, paired ventrolateral and dorsolateral carinae); ventrolateral and median lateral carinae converge distally in segments I–II; intercarinal spaces with abundant weak granulation, except for dorsal surface of segment V which is smooth. Telson almost completely smooth, with few weak granules ventrally and laterally; vesicle deeper and not-too-long as in males of other species of the genus; aculeus long (although shorter than the vesicle) and curved; subaculear tubercle strong and spinoid. Pedipalps: femur with five longitudinal carinae (dorsoexternal, dorsointernal, ventrointernal, ventroexternal and internal median carinae), with abundant granules on all surfaces but ventrally; patella with some well-developed granules on the position of the dorsointernal and ventrointernal carinae, but only two distinct longitudinal carinae (dorsal and median dorsal) formed by aligned granules (Fig. 9); chela acarinate; fixed fingers with six almost linear rows of granules (including the short apical row), the basal is the longest; movable fingers with seven rows. Trichobothriotaxy type A, femur with β configuration [17,18] (Figs. 7–12). Legs: tibia, basitarsus and telotarsus with numerous ventral setae; tibial spur present in legs III–IV; prolateral pedal spur single in legs I–II, bifid in legs III–IV; retrolateral pedal spur present in all the legs.
Male paratype (IAvH-E 100787). Fits well in the description given above, except for the segments count on the pectines: basal lamellae 3:3, middle lamellae 7:8, teeth 17:17. Cheliceral dentition identical to that of the holotype.
Female. Unknown.
Distribution and landscape. Known only from the type locality: northeastern extreme of El Tuparro Natural National Park, Vichada department, at N and W (Fig. 1). According to Villarreal-Leal [19] the landscape occurring at these coordinates, called “Bosque de Cerro”, corresponds to forests and shrubs associated to isolated rocky hills (inselberg) on Igneo-metamorphic complex of the Guyanese shield.
Affinities with Colombian and Venezuelan species. Among all the Colombian species of the genus, A. faguasi sp. nov. seems to be most similar to A. columbiana from northwestern Colombia (Fig. 1), with which it shares the following combination of features that distinguishes both species from all other Ananteris known to occur in the country: and trichobothria are aligned axially (Fig. 12); both est and et trichobothria are located between db and dt (Fig. 11); metasomal segments II and IV possess ten and eight carinae respectively; the chelicerae exhibit a very reticulated pattern on dorsal surface (Fig. 4); pectines have similar number of teeth (A. faguasi sp. nov.: male 17–19; A. columbiana: female 15–19, male 16–18); and the general coloration pattern is similar. The new species can be readily distinguished from A. columbiana since in the former there is no vestige of the median lateral carinae on metasomal segment III, the dorsal surface of metasomal segment V is completely smooth and the other intercarinal spaces on that segment possess only few weak granules, and sternites IV–VI (and VII) are densely granulose throughout (Fig. 6). In contrast, in A. columbiana the median lateral carinae on segment III are present anteriorly, metasomal segment V is densely granulose in all surfaces, and sternite V is completely smooth and VI bears only few granules distally and laterally.
When compared to the Venezuelan species, A. faguasi sp. nov. appears to be most similar to Ananteris asuncionensis González-Sponga, 2006, Ananteris caracensis González-Sponga, 2006, Ananteris norae González-Sponga, 2006 and Ananteris plataensis González-Sponga, 2006, with which it shares the following combination of features: both est and et trichobothria are located between db and dt (Fig. 11; see Figs. 5, 8, 23, 26, 101, 104, 119, 122 in [2]); the metasomal carinal formula is 10:10:8:8:5; the anterior margin of the carapace is not bi-concave (Fig. 5); and pectines have similar number of teeth (A. faguasi sp. nov.: male 17–19; A. asuncionensis: female 17–19; A. caracensis: female 16–18, male 17–18; A. norae: female 16–19; A. plataensis: female 18–20). Of these species, A. plataensis is known from the State of Bolívar in central Venezuela, and the remaining species from the states of Miranda and Nueva Esparta in extreme northern Venezuela (see Map 1 in [2]). Unfortunately, González-Sponga's [2] descriptions are not detailed enough, the features he described are not the same for the different species (making comparisons difficult), and some characters are not appropriately used to distinguish them making the diagnoses rather uninformative (i.e., body size and pectinal teeth number are used to distinguish species even when differences can be explained in means of normal intraspecific variability). This way, that author did not provide information regarding the relative position of the ventral trichobothria on pedipalp chela for any of the species, and, among the four species mentioned above, he only described the coloration of the dorsal surface of the chelicerae for A. asuncionensis. Therefore, so far it is impossible to know if A. faguasi sp. nov. differs from these other species in these two characters, widely used through this paper. Despite this situation, the new species differs from its apparently most similar species from the neighboring country in having the sternites IV–VII densely granulose (Fig. 6), whereas these are coriaceous in A. asuncionensis, A. caracensis and A. norae, and have no ornamentation in A. plataensis (except VII which scarcely granulose) [2]. Also, it differs from A. asuncionensis in having the median ocular carinae granulose (Fig. 5) and conspicuous median lateral carinae on metasomal segments I–II, whereas in A. asuncionensis the median ocular carinae are smooth and the median lateral carinae on segments I–II are vestigial [2]. Finally, A. faguasi sp. nov. can be also distinguished from A. plataensis since in the later species the intermedian carinae are vestigial on metasomal segment II [2].
Ananteris volschenki sp. nov.

Ananteris volschenki sp. nov. 13–14. Male holotype (IAvH-E 100788), dorsal and ventral views. 15–16. Female paratype (IAvH-E 100789), dorsal and ventral views. Scale bars equal 5 mm. (For interpretation of the references to colour in this figure, the reader is referred to the web version of this article.)

Ananteris volschenki sp. nov., male holotype (IAvH-E 100788). 17. Dorsal view of left chelicera. 18. Anterior margin of the carapace (only granulation of the median ocular carinae is included). 19. Sternite VII. 20–25. Distribution of the trichobothria (illustrated from the right pedipalp). 20. Femur, dorsoexternal view. 21. Femur, internal view. 22. Patella, dorsal view. 23. Patella, external view. 24. Chela, external view. 25. Chela, ventral view. SC = Submedian Carina. Scale bars equal 0.5 mm.
Type material.Holotype: COLOMBIA: Vichada department: adult male from Cumaribo, Santa Rita, Parque Nacional Natural El Tuparro, N, W, 135 m (210 m sensu Villareal-Leal [19]), Mata de Monte, pitfall, 08–10 February 2004, I. Quintero and E. González (IAvH-E 100788).
Paratype: COLOMBIA: Vichada department: one adult female from Cumaribo, Santa Rita, Parque Nacional Natural El Tuparro, N, W, 135 m (210 m sensu Villareal-Leal [19]), Mata de Monte, Winkler, 08 February 2004, I. Quintero and E. González (IAvH-E 100789).
Non-type material. COLOMBIA: Vichada department: one juvenile (presumably male) with identical collection data to that of the holotype (IAvH-E 100790).
Etymology. Patronym dedicated to Erich S. Volschenk, scorpiologist at the Department of Terrestrial Invertebrates of the Western Australian Museum (Perth), in recognition of his contributions to scorpiology and acknowledgment of his advice in the author's research.
Diagnosis.Ananteris volschenki sp. nov. differs from all other species of the genus by the following unique combination of features: and trichobothria are not aligned axially but is located on an external position in relation to (Fig. 25); only et trichobothrium is located between db and dt on pedipalp fixed finger, with est considerably basal to db (Fig. 24); the anterior margin of the carapace is bi-concave (Fig. 18); the median ocular carinae are granulose (Fig. 18); the metasomal carinal formula is 10:8:8:8:5 with all carinae complete; the dorsal surface of chelicerae exhibits a complete and very reticulated pattern (Fig. 17); sternite VII presents incomplete submedian carinae (Fig. 19); pectines have 15–16 teeth (female 15–16, male 15–15); and sternite III is smooth, IV bears few weak granules laterally, and V–VII have abundant granulation throughout.
Description of the male holotype (IAvH-E 100788).
Coloration. General coloration yellowish with variegated pigmentation over almost the entire body (except ventrally) and appendages. Carapace predominantly yellow with some brown spots and bands; anterior margin yellow, posterior margin brown; a brown triangle pointing anteriorly is present in front of the median ocular tubercle, which is black. Chelicerae with coxa yellow; hand with a complete reticular pattern on dorsal surface; movable finger with a large dorsal brown area and reddish teeth; fixed finger yellow with reddish teeth (Fig. 17). Coxapophyses I–II with brown spots anteriorly; coxae I–IV, sternum, genital operculum, pectinal basal piece, pectines and sternites III–V completely yellow, without marks; sternite VI with few brown spots laterally; sternite VII with inconspicuous brown areas between the midline and the lateral margins; sternite III with a narrow transverse posterior median hyaline area. Tergites I–VI predominantly yellow anteriorly, brown posteriorly, with a yellow longitudinal area at each side of the midline; each side of tergites I–VI with two transverse yellow lines converging near the longitudinal lines, arrow-like; tergite VII with brown mottling laterally, predominantly yellow dorsally but with a brown area on the place of the axial carina; lateral margins of tergites I–VII yellow. Metasoma predominantly yellowish; dorsal intercarinal spaces of segments I–III predominantly yellow, brown-spotted medially; segments I–V with variegated pigmentation; segment IV reddish; V dark-red, lighter anteriorly. Telson reddish, lighter than segment V, with inconspicuous brownish spots on the distal section of the vesicle; aculeus yellowish basally; subaculear tubercle yellow. Pedipalps predominantly yellow; coxa and trochanter with brown regions; femur with variegated pigmentation in dorsal, internal and external surfaces, predominantly yellow ventrally except for a distal brown area, insertion of all the trichobothria yellow; patella with variegated pigmentation in all surfaces, predominantly yellow medially and brownish in the extremes, insertion of all the trichobothria yellow; hand brown in all surfaces, with some light-brown areas; fixed and movable fingers predominantly yellow, except basally where brown, slightly brownish in the distal half. Legs with variegated pigmentation, brown spots on all segments except for the telotarsus that is completely yellow.
Morphology. Measurements in Table 1. Carapace: densely covered with rounded granules, especially on the brown regions; lateral margins not parallel; anterior margin with two slight concavities (Fig. 18); median ocular carinae evident, others inconspicuous; a median depression anterior to the ocular tubercle and a posterior median longitudinal furrow are evident; median ocular tubercle low, located on the posterior portion of the anterior third of the carapace, with abundant granulation on the median ocular carinae (Fig. 18); lateral ocular tubercles each with three ocelli (presumably four). Chelicerae: with abundant setae on the internal and ventral surfaces; cheliceral dentition characteristic of the family Buthidae [20]. Movable finger externally with two small basal teeth, one median pronounced, one subdistal slightly shorter than the median, and one distal tooth. Internally with two well-developed teeth, one basal somewhat rounded and one median pronounced, and one distal tooth. Fixed finger externally with one basal and one median tooth mounted onto a bicuspid, one subdistal, and one distal tooth. Internally with only one pronounced tooth located slightly basal in respect to the external subdistal. Coxosternal region: with abundant setae; sternum subtriangular, with a deep median depression; coxae I and coxapophyses I–II smooth; coxae II–IV with abundant minute granulation; coxapophyses I–II with dense pilosity anteriorly. Genital operculum and pectines: genital operculum divided longitudinally; pectinal basal piece as long as wide; pectines long, surpassing the lateral margins of sternite III (Fig. 14); segments count on the pectines: basal lamellae 3:3, middle lamellae 6:7, teeth 15:15, fulcra absent. Sternites: III–VII densely covered with long fine translucent setae; sternite III smooth; IV with few weak granules laterally; V–VII with abundant granulation throughout, greater in size on sternite VII; VII with parallel submedian carinae, evident and incomplete, present in the two most distal thirds of the sternite, and lateral carinae completely absent (Fig. 19); spiracles oval elongate. Tergites: with similar granulation to that of the carapace; axial carina only evident in the posterior half of tergites I–VI; dorsolateral carinae absent; tergite VII tetracarinate (paired dorsolateral and lateral carinae, incomplete), a median elevation and some slightly greater granules are present on the position of the axial carina. Metasoma: with abundant setae; all carinae complete; segment I with ten carinae (paired ventrosubmedian, ventrolateral, median lateral, dorsolateral and dorsosubmedian carinae); segments II–IV with eight (median lateral carinae absent); segment V with five (ventromedian, paired ventrolateral and dorsolateral carinae); median lateral carinae converge distally towards the ventrolateral in segment I; all carinae serrulose, except for the dorsolateral on segment V which are smooth in the distal half; intercarinal spaces with few weak granulation, except for dorsal surface of segment V which is smooth. Telson completely smooth; vesicle long; aculeus long (although shorter than the vesicle) and curved; subaculear tubercle strong and spinoid. Pedipalps: femur with five longitudinal carinae (dorsoexternal, dorsointernal, ventrointernal, ventroexternal and internal median carinae), with few granules on the intercarinal spaces; patella with some well-developed granules on the position of the dorsointernal and ventrointernal carinae, but only two clearly distinct longitudinal carinae (dorsal and median dorsal) formed by aligned granules (Fig. 22); chela acarinate, but with four conspicuous granules arranged in two diads on the internal surface; fixed fingers with six almost linear rows of granules (including the short apical row), the basal is the longest; movable fingers with seven rows. Trichobothriotaxy type A, femur with β configuration [17,18] (Figs. 20–25). Legs: tibia, basitarsus and telotarsus with numerous ventral setae; tibial spur present in legs III–IV; prolateral pedal spur single in legs I–II, bifid in legs III–IV; retrolateral pedal spur present in all the legs.
Female paratype (IAvH-E 100789). Fits well with the description given above, except for the following: Coloration: metasomal segments predominantly yellow, without red; telson yellow, with the aculeus reddish (Figs. 15–16). Morphology: greater in size (see morphometric measurements); pectines smaller, with shorter teeth (Figs. 15–16); segments count on the pectines: basal lamellae 3:3, middle lamellae 7:7, teeth 15:16. Cheliceral dentition identical to that of the holotype.
Distribution and landscape. Known only from the type locality: northeastern extreme of El Tuparro Natural National Park, Vichada department, at N and between W and W (Fig. 1). According to Villarreal-Leal [19] the landscape occurring at these coordinates corresponds to isolated forests, called “Matas de Monte”, over well-drained plains with Eolic sands cover.
Affinities with Colombian and Venezuelan species. Among all the Colombian species of the genus, A. volschenki sp. nov. seems to be most similar to A. myriamae from central Colombia (Fig. 1), with which it shares the following combination of features that distinguishes both species from all other Ananteris known to occur in the country: and trichobothria are not aligned axially but is located on an external position in relation to (Fig. 25; see Fig. 13 in [5]); the chelicerae exhibit a very reticulated pattern on dorsal surface (Fig. 17; see Fig. 3 in [5]); pectines have similar number of teeth (A. volschenki sp. nov.: female 15–16, male 15–15; A. myriamae: female 15–16); the anterior margin of the carapace is bi-concave (Fig. 18; see Fig. 6 in [5]); and sternite VII possesses vestigial submedian carinae (Fig. 19). The new species can be readily distinguished from A. myriamae since in the former the metasomal carinal formula is 10:8:8:8:5, and est trichobothrium is considerably basal to db on pedipalp fixed finger (Fig. 24). In contrast, in A myriamae the metasomal carinal formula is 10:10:10:6:5, and est trichobothrium is located beside db (see Fig. 12 in [5]). In addition, the coloration pattern is different, especially on pedipalp chelae: in the new species the hand is brown and the fingers predominantly yellow (Figs. 13–16), whereas in A. myriamae the hand is completely yellow and the fingers brown (see Fig. 2 in [5]).
When compared to the Venezuelan species, A. volschenki sp. nov. appears to be most similar to Ananteris venezuelensis González-Sponga, 1972 from eastern Venezuela in the State of Bolívar (see Map 1 in [2]), with which it shares that only et trichobothrium is located between db and dt on pedipalp fixed finger (Fig. 24; see Fig. 170 in [2], Fig. 4b in [21]), and the metasomal carinal formula 10:8:8:8:5. The new species can be readily distinguished from A. venezuelensis since the ventral trichobothria of the pedipalp chela are markedly unaligned axially in the former species (Fig. 25) but aligned in the later (see Fig. 4a in [21]).
4 Remarks
Even though the coordinates at which each of the new species described in the present paper are very close together (Fig. 1), no evidence exists to indicate that these are sympatric. Taking into account that the two unique specimens available of A. faguasi sp. nov. were collected at the same landscape, namely “Bosque de Cerro”, whereas the three specimens of A. volschenki sp. nov. were captured at “Mata de Monte”, it is likely that this may be due to habitat specialization. Further studies at El Tuparro Natural National Park would provide additional information on this matter.
5 Other material examined
In the course of this study, a number of specimens of Ananteris were examined, apart from the two new species. These specimens are detailed bellow.
Ananteris arcadioi Botero-Trujillo, 2008: COLOMBIA: Meta Department: male holotype, Puerto Gaitán, Altamira, Club Los Llaneros, 140 m, into forest, ad hoc, at night, 19 October 2006, I. Gélvez (MPUJ-SCO-356).
Ananteris columbiana Lourenço, 1991: COLOMBIA: Atlántico Department: 1 female, Puerto Colombia, El Nisperal, 100 m, 10–15 June 2006, G. Fagua (MPUJ-SCO-334). Bolívar Department: 1 female, Cartagena, Isla Barú, 40 m, pitfall, 14 October 2006 (MPUJ-SCO-358); 1 male, Zambrano, Hacienda Monterrey, 70 m, 01 December 1997, F. Fernandez and G. Ulloa (ICN-MHN-As-446); 1 juvenile, Zambrano, Hacienda Monterrey, 120 m, dry forest, pitfall, August 1996, F. Escobar (ICN-MHN-As-176); 1 female, Zambrano, Hacienda Monterrey, 70 m, 19 October 1993, F. Fernandez (ICN-MHN-As-595); 1 female, Zambrano, 20 m, 25 August 1992, A. Molano (ICN-MHN-As-111); 1 juvenile, Santa Catalina, Hacienda El Ceibal, 20 m, October 1999, E. Flórez and Biology students (ICN-MHN-As-294); 1 female, Parque Nacional Natural Colorados, La Yaya, 280 m, N, W, pitfall, 24–26 May 2001, E. Deulufeut, M. 1731 (IAvH-E 100770); 1 female, Parque Nacional Natural Colorados, Alto El Mirador, 400 m, N, W, pitfall, 05–09 July 2001, E. Deulufeut, M. 1956 (IAvH-E 100771); 1 female, Parque Nacional Natural Colorados, Alto El Mirador, 400m, N, W, pitfall, 02–06 December 2001, E. Deulufeut, M. 2644 (IAvH-E 100772); 1 female, Parque Nacional Natural Colorados, La Suiris, 126 m, N, W, pitfall, 18–21 November 2000, E. Deulufeut, M. 937 (IAvH-E 100773); 1 female, Parque Nacional Natural Colorados, La Suiris, 126 m, N, W, pitfall, 18–20 December 2000, E. Deulufeut, M. 976 (IAvH-E 100774); 1 female, Parque Nacional Natural Colorados, Villa Roca, 180 m, N, W, pitfall, 19–22 August 2001, E. Deulufeut, M. 2055 (IAvH-E 100775); 1 juvenile [male (?)], Parque Nacional Natural Colorados, Villa Roca, 180 m, N, W, pitfall, 24–26 May 2001, E. Deulufeut, M. 1729 (IAvH-E 100776). Córdoba Department: 1 male, Pueblo Nuevo, Hacienda Toronto, 30 m, 07–13 June 2004, J.D. Lynch (ICN-MHN-As-585). Magdalena Department: 1 male and 1 female, Santa Marta, Parque Nacional Natural Sierra Nevada de Santa Marta, 120 m, December 2006, J.A. Noriega (MPUJ-SCO-363, 364); 1 female, Santa Marta, near Quebrada Minca, 2 m, November 1976 (ICN-MHN-As-121); 1 female, Parque Nacional Natural Tayrona, Zaino, 50 m, N, W, pitfall, 04–06 December 2000, H. Henriquez, M. 1013 (IAvH-E 100777); 1 female, Parque Nacional Natural Tayrona, Pueblito, 225 m, N, W, pitfall, 30 September 2000, H. Henriquez, M. 660 (IAvH-E 100778); 2 females, Parque Nacional Natural Tayrona, Pueblito, 225 m, N, W, pitfall, 29 July 2000, H. Henriquez (IAvH-E 100779,100780); 1 juvenile [male (?)], Parque Nacional Natural Tayrona, Pueblito, 225 m, N, W, pitfall, 15 August 2000, H. Henriquez (IAvH-E 100781).
Ananteris dorae Botero-Trujillo, 2008: COLOMBIA: Nariño Department: female holotype, Reserva Natural La Planada, permanent plot, N, W, 1885 m, pitfall, 02–04 May 2001, G. Oliva, M. 2369 (IAvH-E 100763).
Ananteris ehrlichi Lourenço, 1994: COLOMBIA: Caquetá Department: 1 female, La Montañita, Santuario Las Iglesias, Itarca, 330 m, 25 April 2004, M. Agudelo (ICN-MHN-As-579); 1 female, Parque Nacional Natural Chiribiquete, Río Mesay, 20 January 2000, F. Quevedo (ICN-MHN-As-361); 1 female, Parque Nacional Natural Chiribiquete, Río Sararamano, April 2000, F. Quevedo (ICN-MHN-As-342).
Ananteris gorgonae Lourenço and Flórez, 1989: COLOMBIA: Cauca Department: 1 male, Parque Nacional Natural Gorgona, El Mirador, 180 m, pitfall, 03–04 February 2001, R. Duque (ICN-MHN-As-427); 1 male, Parque Nacional Natural Gorgona, El Helechal, 30 m, N, W, pitfall, 17–19 July 2001, H. Torres, M. 2003 (IAvH-E 100766); 1 male, Parque Nacional Natural Gorgona, Alto El Mirador, 180 m, N, W, pitfall, 18–20 January 2001, H. Torres, M. 1245 (IAvH-E 100767); 1 male, Parque Nacional Natural Gorgona, El Helechal, 30 m, N, W, pitfall, 08–09 March 2002, H. Torres, M. 3098 (IAvH-E 100768); 1 adult [gynandromorph (?)], Parque Nacional Natural Gorgona, El Roble, 130 m, N, W, pitfall, 20–21 February 2001, H. Torres, M. 1369 (IAvH-E 100769).
Ananteris aff. gorgonae: COLOMBIA: Valle del Cauca Department: 1 female, Buenaventura, Bahía de Malaga, Base Naval, 5 m, April 1989, L.A. Millan (ICN-MHN-As-391).
Ananteris leilae Lourenço, 1999: COLOMBIA: Chocó Department: 1 male, Acandí, Capurganá, Los Ríos, 230 m, 25 April 2007, M. Gutierrez (MPUJ-SCO-374); 1 female, Acandí, Capurganá, Jardín Botánico del Darién, 40 m, Rastrojo, 11 October 2007, C. Acosta, J. Alfonso and C. Cocoma (MPUJ-SCO-377).
Ananteris myriamae Botero-Trujillo, 2007: COLOMBIA: Meta Department: female holotype, Villavicencio, Vereda El Carmen, 850–1000 m, into forest, pitfall, 23 December 2005, M. Viola (MPUJ-SCO-245); female paratype, Villavicencio, Vereda El Carmen, 850–1000 m, Río Caño Blanco, under litter, ad hoc, at night, 18 April 2005, R. Botero-Trujillo (MPUJ-SCO-039).
Acknowledgements
The author is most grateful to an anonymous reviewer for his valuable comments that led to the improvement of the manuscript. Special thanks are due to Diego Perico, Monica Ospina and Edwin Torres (Instituto de Investigación de Recursos Biológicos Alexander von Humboldt, Villa de Leyva, Colombia) for arranging the loan of some of the specimens examined for this contribution (including the type material), to Eduardo Flórez and Carlos Sarmiento (Instituto de Ciencias Naturales, Bogotá) for permission to visit the scorpion collection of the ICN-MHN, and to Luis E. Franco (IAvH) for his help in logistic matters. E. Flórez kindly loaned his personal stereoscope for obtaining the measurements. The following persons assisted in acquiring relevant literature: Fernando J.M. Rojas-Runjaic (Museo de Historia Natural La Salle, Venezuela), Oscar F. Francke and Carlos E. Santibañez (Universidad Nacional Autónoma de México), E. Flórez (Instituto de Ciencias Naturales, Bogotá), Mónica Higuera (Pontificia Universidad Javeriana, Bogotá), and František Kovařík (Praha, Czech Republic). This study was performed at the Laboratorio de Entomología of the Pontificia Universidad Javeriana (Bogotá); the author is grateful to its director, Giovanny Fagua, for providing space for work and access to laboratory equipment and material. Finally, thanks also go to Miguel León and Nestor García (PUJ) for the loan of the camera lucida.