

1 Introduction
The Philippines can be regarded as a hotspot of phasmatodean biodiversity. So far, approximately 6.7% of the world phasmatodean fauna has been described or recorded from the archipelago [1], representing several major lineages of the clade Euphasmatodea sensu Bradler [2] from Asia and Australasia.
Aschiphasmatidae form a distinctive group of relatively small-sized extant euphasmatodeans, comprising 15 genera and 26 species distributed from southern India and southern China to Southeast Asia [1]. Members of this taxon live in lowland and montane dipterocarp forests, where they can be found from within the low-growing vegetation, up to the canopy level, several meters above ground. In these habitats, they express varying degrees of wing reduction, including macropterous, brachypterous, micropterous, and apterous taxa. However, when the fore wing is present, it is always highly simplified, and develops in the form of a small filament or a minute longitudinally curved lobe. When threatened, these stick insects can jump backwards and secrete a defensive spray from the prothoracic exocrine glands. Females drop lentil-shaped eggs, which in certain taxa are equipped with peculiar setae-like structures that may function as attachment devices [3].
Tilgner [4] based on a cladistic analysis of morphological characters of the adult female and egg capsule, pointed out that Aschiphasmatidae may form a monophyletic lineage and the sister group of all remaining euphasmatodeans. Phylogenetic analyses of mitochondrial COI and COII, along with nuclear 28S rRNA and histone H3 genes sequence data by Buckley et al. [5] suggested a clade comprising the Afrotropical genus Xylica Karsch, 1898 and the Aschiphasmatidae, representing the sister group of all remaining lineages of Euphasmatodea. A basal placement, and a close relationship between Xylica and Aschiphasmatidae was also supported in a recent cladistic study by Bradler [6]. From the standpoint of intraphyletic relationships, the lineage is currently divided into two groups, whose distinction is traditionally based upon the presence (Aschiphasmatini) or absence (Dajacini) of denticulations on the pretarsal claws [3]. In addition, putative autapomorphies substantiating the validity of these two groups have been proposed by Bradler [6].
Present knowledge of Philippine Aschiphasmatidae is scanty. Gottardo [7] summarized the few published reports, recording four genera and five species from the islands of Luzon, Mindanao, Palawan, and Panay. During the work on ongoing project “Systematics of Philippine Phasmatodea” [7–11], the author examined new samples of micropterous Aschiphasmatidae characterized by a set of peculiar apomorphic features which allowed recognition of a new genus and species. Also, the specimens of the new taxon were collected on Mindoro island, where members of this group were not yet reported.
The aim of this article is to provide a characterization and description of this new taxon, based on the study of adult specimens of both sexes. Specifically, light microscopy (LM) was used to observe the general external features of the body, and scanning electron microscopy (SEM) was used to characterize some aspects of the morphology of the legs. The findings resulting from the morphological study showed some novelties with potential implications for the systematics of the Phasmatodea and Aschiphasmatidae, and were discussed in a phylogenetic context.
2 Materials and methods
The specimens employed in this study were dried and pinned. Photographs of the habitus were taken with a Nikon D50 digital camera. Observations on the gross morphology were performed using a Zeiss Stemi DV4 stereo light microscope, and drawings were prepared with the aid of a camera lucida. Mid legs were prepared for SEM. The portion of the leg was quickly cleaned in 5% KOH, air-dried in a stove at 60 °C, mounted on an aluminium stub using double stick adhesive, gold-coated in a Balzers MED 010 sputtering device, and observed with a Philips XL20 scanning electron microscope operated at 10 kV. The type specimens were lodged at the Zoologische Staatssammlung München, Germany (ZSM), and in the personal collection of Oskar V. Conle, Bolsterlang, Germany (OC, affiliated to the collection of the ZSM). The taxon here described represents a new monotypic genus. For this reason the description of the genus and species are equivalent. In the results section of the paper, the generic diagnosis only summarizes the most significant features, whilst a full and detailed description is provided for the species. In this study the arrangement of Aschiphasmatidae follows the taxonomy of Bragg [3,12].
3 Results
Order PHASMATODEA Jacobson & Bianchi, 1902
Family ASCHIPHASMATIDAE Brunner von Wattenwyl, 1893
Subfamily ASCHIPHASMATINAE Brunner von Wattenwyl, 1893
Tribe ASCHIPHASMATINI Brunner von Wattenwyl, 1893
Genus Dallaiphasma gen. n.
3.1 Diagnosis of the new genus
The new genus is characterized by the structure of the vertex, which is strongly raised, distinctly surpassing the pronotum in height, and notably cone-shaped. As a consequence of this peculiar head structure, the chewing mouthparts are directed slightly downward. The tegmina is not developed; the hind wing is lobiform, lateral. The whole of dorsal and lateral surfaces of femora is swollen and rounded, lacking dorsal carinae; the ventro-anterior and ventroposterior carinae are distinct. The tibiae are rounded in cross-section, without any carinae. An area apicalis is well developed on all tibiae; it is cup-shaped, and features a depressed membranous posterior lateral region, and a strongly sclerotized central apical region. The pretarsal claws bear minute denticulations on the two ventral longitudinal keels. The abdominal tergum I is broader than long, with well-defined margins. In the male the genital region features a sharply triangular sternite IX, a vomer with its distal half strongly curved upward, and a single row of toothlike projections on the ventroposterior margin of the tergum X. The female genital region exhibits a boat-shaped sternum VIII, and a short ovipositor which is completely hidden by the sternum VIII.
Etymology: the new genus-group name is a patronymic dedicated to the author's teacher Professor Romano Dallai (University of Siena), to acknowledge for his continued support during the author's PhD studies in evolutionary zoology, and also in recognition of his significant contributions to the knowledge of the Arthropoda. The Greek word “Phasma” (meaning, ghost) is a classical genus-group names ending in stick insects.
Type species: Dallaiphasma eximius sp. n., by present designation.
3.2 Description of the new species
Dallaiphasma eximius sp. n. (Figs. 1–5; Table 1).

Habitus photographs of Dallaiphasma eximius gen. et sp. n. in lateral view. A. Holotype male. B. Paratype female.

Scanning electron micrographs of the mid femora of Dallaiphasma eximius gen. et sp. n. in lateral view. A. Paratype female, the ventral femoral carinae are unarmed. B. Paratype male, the ventral femoral carinae are equipped with well-developed spines. C. Detailed view of a spine on the ventro-anterior carina of paratype male. T: trochanter; F: femur; VAC: ventro-anterior carina; S: spine.

Scanning electron micrographs of the mid tibia of Dallaiphasma eximius gen. et sp. n. in ventral view. A. Whole of the area apicalis of paratype male. B. Detailed view of the sample shown in A, the cuticular surface exhibits structural differences in the two regions of the area apicalis. CF: cuticular fold; MR: membranous posterior lateral region; SR: sclerotized central apical region.

Scanning electron micrographs of mid tarsus of Dallaiphasma eximius gen. et sp. n., paratype male. A. Tarsomere IV and pretarsus in ventral view. B. Detailed view of the outer margin of the arolium in ventral view. C. Pretarsal claws in lateral view, the arrowheads show the presence of minute denticulations on the ventral lateral keel. D. Microstructure of the euplantula. AR: arolium; CL: claw; EU: euplantula; TA4: Tarsomere IV; TA5: pretarsus; UT: unguitractor plate.
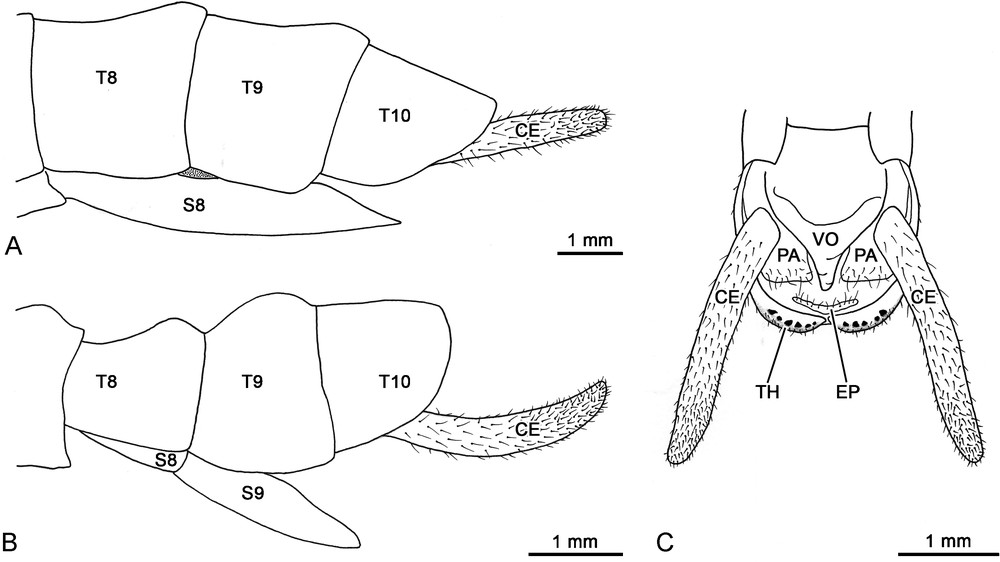
Schematic drawings of Dallaiphasma eximius gen. et sp. n. terminalia. A. Paratype female, lateral view. B. Paratype male, lateral view. C. Paratype male, ventral view. CE: cercus; EP: epiproct; PA: paraproct; S8-S9: abdominal sternites 8-9; T8-T10: abdominal tergites 8-10; TH: thorns; VO: vomer.
Morphometric data for the type specimens of Dallaiphasma eximius gen. et sp. n.
| Parameter | ♂ holotype | ♂ paratype | ♀ Paratype |
| (ZSM) | (OC) | (OC) | |
| Length (mm) | |||
| Body | 28.8 | 30.4 | 42.0 |
| Antenna | 19.8 | - | - |
| Head | 2.3 | 2.5 | 3.2 |
| Pronotum (PL) | 2.6 | 3.1 | 3.9 |
| Mesonotum (MOL) | 3.5 | 4.0 | 5.9 |
| Metanotum (MAL) | 1.5 | 1.6 | 2.9 |
| Median segment (MSL) | 2.3 | 2.1 | 2.9 |
| Hind wing | 3.7 | 4.2 | 2.5 |
| Cercus | 2.6 | 2.7 | 3.3 |
| Fore femur | 4.7 | 4.7 | 5.6 |
| Fore tibia | 4.3 | 4.5 | 5.7 |
| Mid femur | 4.1 | 4.2 | 4.8 |
| Mid tibia | 4.2 | 4.5 | 5.4 |
| Hind femur | 5.9 | 6.1 | 7.3 |
| Hind tibia | 6.5 | 6.6 | 8.3 |
| Morphometric ratios | |||
| MOL divided by PL | 1.35 | 1.29 | 1.51 |
| MAL divided by MOL | 0.43 | 0.40 | 0.49 |
| MSL divided by MAL | 1.53 | 1.31 | 1.00 |
Type material: holotype, male, Philippines, Mindoro Island, Mt. Halcon, VIII.2006 (depository: ZSM). Paratypes, one male, one female, same data as the holotype (coll. OC).
Etymology: the specific name “eximius” (from Latin, extraordinary) refers to the distinctiveness of the new taxon.
Diagnosis and general description: this new species is one of the smallest representatives of Aschiphasmatidae: males are approximately 30 mm, females 40 mm. In both sexes the dorsal and lateral surfaces of the body, and the legs are strongly shiny, whilst the ventral surface of body is only slightly glossy. The integument is rather smooth; setation is developed mainly on the ventral surfaces of the body and legs. The head and femora are vivid black. The compound eyes are mid brown. The antennae, tibiae, tarsi, thorax and abdomen are deep reddish-brown. The hind wings are yellowish. The habitus of the male and female is illustrated in Fig. 1. A full set of measurements and morphometric ratios is given in Table 1.
Male morphology: the head capsule is wider than long. The dorsal surface of head features an oblong depression between the antennal sockets, and a slight median longitudinal coronal suture along the vertex. The gena is approximately as broad as the greatest diameter of the compound eye. As typical of the majority of adult Aschiphasmatidae, the antenna is slightly shorter than body length. The holotype exhibits a complete left antenna, composed of 29 antennomeres. The scape is rectangular and thickened from base to apex. The pedicel is cylindrical, only slightly shorter than scape. The first flagellomere is more narrower and shorter than pedicel. The antennal flagellum is densely covered with short sensilla and setae. The compound eye is circular, and protrudes hemispherically. Ocelli are lacking. The maxillary palpus is pentamerous, with rather cylindrical articles. The lacinia exhibits only two apical teeth. The labial palpus is trimerous, with moderately flattened articles. The ventral surface of the head features a small median gular sclerite. The prothorax is slightly longer than head, and wider than long. The sub-trapezoidal pronotum (Fig. 1A) is characterized by a marked transverse depression approximately on the anterior third. The portion of pronotum in front of the depression is slightly swollen, and features a concave front margin, and a well-differentiated glandular area at the level of the two anterolateral corners. Each pronotal glandular area consists of a bean-like impression equipped with a slot-like opening of the prothoracic exocrine gland. The portion of pronotum behind the depression is raised to form a hump, which is higher than anterior portion; the posterior margin of pronotum is almost straight. The pronotal sides (or paranota) are distinctly developed laterally, and slightly cover the pleural region. The prothoracic coxopleurite is rather cuneiform-shaped. The prothoracic basisternite is sub-trapezoidal. The mesothorax (Fig. 1A) widens evenly from the anterior to the posterior end; the posterior is slightly more than one and a half times wider than anterior. The mesonotum is tectiform and increasingly raised towards the posterior area; its hind margin is slightly rounded and bears a prominent median mound, representing the higher point of the dorsal body surface. The mesothoracic episternum is elongated, whilst the epimeron is small and sub-triangular; the two sclerites are separated by a distinct pleural suture. The mesothoracic basisternite is elongated and bears a median keel; the furcasternite is much shorter than the basisternite. The metathorax (Fig. 1A), which also incorporates the first abdominal segment, is nearly parallel-sided, and of equal length as the mesothorax. The metanotum is distinctly shorter than mesonotum, and wider than long; much of its surface is occupied by a large rounded tubercle, which raises medioposteriorly. The hind margin of the metanotum is well differentiated, which allows one to easily distinguish the boundary with the first abdominal tergum (or median segment). The metathoracic pleural region is basically structured as in the mesothorax. The metathoracic basisternite features a very short median keel on the anterior portion; the furcasternite is only slightly shorter than the basisternite. The tegmina are lacking. The hind wing (Fig. 1A) consists of a small lateral lobe without any distinct venation; its shape is scalene triangular, and the posterior end extends moderately beyond the first abdominal tergum. The legs (Fig. 1A) are short and stout; the hind leg only slightly projects beyond the ninth abdominal tergum. The coxae are unarmed. Trochanters are very short and fused to femora, as in all other euphasmatodean taxa. The fore femur is evenly straight and basally uncompressed; the ventro-anterior carina shows a row of seven distinct spines, whilst the ventroposterior carina possesses 2–5 small distal spines. The mid femur (Fig. 2B) is armed with 11–14 prominent spines along the ventro-anterior carina, and only three-four small distal spines on the ventroposterior carina. Likewise, the hind femur bears 15–17 prominent spines along the ventro-anterior carina, and only 4–7 smaller spines on the ventroposterior carina. The spines (Fig. 2C) are stout, angularly oriented towards the distal direction, and show a prominent gibbosity near the distal end, which abruptly tapers to form a sharply rounded apex. The size of the spines is larger in the central portion of the carina, and gradually decreases towards the proximal and distal ends. Every tibia is slightly swollen at the distal apex, where an area apicalis is developed (Fig. 3A). This is represented by a cup-like depression, equipped with a plate-like element which is located centro-apically. The cuticle of the area apicalis is unevenly sclerotized. In the depressed posterolateral region the cuticle is flexible, weakly sclerotized, forms cuticular folds, and lacks the setation and fine ornamentation that characterize the more sclerotized areas of the leg (Fig. 3B). This region surrounds the plate-like element, which is convex and features a medially pointed distal margin. Contrarily to the posterolateral region, the plate-like element is strongly scerotized, and show both well-developed setation and fine cuticular ornamentation. The five-segmented tarsus is slightly more than half the length of the corresponding tibia. The basitarsus is shorter than combined length of tarsomeres II-IV; tarsomeres I-IV are progressively shorter. The pretarsus of fore and hind tarsi is shorter than combined length of tarsomeres I-IV; it is longer in the case of mid tarsi. The pretarsus regularly widens towards the distal end, as typical of all other phasmatodeans. Its ventral surface (Fig. 4A) shows a distinct median longitudinal carina, and an indented distal margin which bears small acanthae; an euplantula is not developed. The unguitractor plate is well developed (Fig. 4A). The arolium is rounded and broad (Fig. 4A). Its adhesive surface is smooth, but granule-like outgrowths are distinctly developed along the region which faces the outer margin (Fig. 4B). The pretarsal claws are identical in shape and size. The distal third of the claw is strongly curved and features two ventral lateral keels. The basal portion of each keel is differentiated into a row of 5–6 minute denticles (Fig. 4C). The ventral surface of tarsomeres I-IV features a short median carina proximally, and a well-developed oblong euplantula distally. The membranous surface of the euplantula (Fig. 4D) is smooth, and its microstructure consists of tightly packed pentagons and hexagons; each polygon is approximately 5 microns wide. The first abdominal sternum is fused with the metasternite, as occurs in all other phasmatodeans. The first abdominal tergum is well defined and easily distinguishable from the metanotum. It is much wider than long, slightly longer than the metanotum, and bears a small posteromedian mound. The remaining abdominal segments are rather short: terga II-III are slightly broader than long; terga IV-VII become longer than broad; terga VII-X are also much broader than long. The posterior margin of terga II-IX is swollen dorsally, and slightly raised than the anterior margin. The tergum X (Fig. 5B) is only slightly longer than tergum IX; it is tectiform in structure, particularly in the posterior end. The hind margin of tergum X is rounded, with a weakly developed median notch represented by a small area of thinner surface. The median notch divides the hind margin into two more thickened portions, each armed ventrally with a row of 5–6 toothlike projections (Fig. 5C). These external processes, also known as thorn pads, are stout, strongly sclerotized, and project ventrally. The sternum IX (or male subgenital plate) is twice as long as sternum VIII. It is a triangular undivided plate, which gradually narrows from the base to the pointed apex, and slightly extends over the tergum IX. The triangular vomer is broad at base, and increasingly tapers towards the apex; its distal half is strongly curved upward. Paraprocts are short, with a rather straight posterior margin, and are mainly concealed by the cerci. The epiproct is very short, and does not extend beyond the hind margin of tergum X (Fig. 5C). The cercus is strongly elongated, about twice as long as tergum X, and very gently incurved; it is more or less cylindrical, covered with numerous erect to suberect sensilla and setae, and its posterior part gradually tapers to a rather rounded tip which lacks any spine (Fig. 5B–C).
Female morphology is as described for the male, except as follows (Fig. 1B): the transverse depression of the pronotum is weakly developed, and both the anterior and posterior areas are moderately raised. The hind margin of the mesonotum lacks a distinct median mound. The metanotum bears only a faint posteromedian hump. The lobiform hind wing is rather heart-shaped, and extends moderately beyond the hind margin of the metanotum. The femora are less robust than those of the male, and the ventral carinae are characterized by the complete absence of any spination. The first abdominal tergum is as long as the metanotum, lacks a distinct posteromedian mound, and is not laterally concealed by the hind wings. The posterior margin of terga II-IX is very moderately swollen dorsally. The tergum X (Fig. 5A) is shorter than tergum IX, and shows an evenly rounded, unmodified hind margin. The sternum VIII (the female subgenital plate) is rather short, about one and a half times as long as sternum VII. It is boat-shaped and lacks a median keel, with an acutely pointed hind margin which reaches half length of tergum X. The cercus is about one and a half times the length of tergum X (Fig. 5A).
Remarks: the egg of D. eximius gen. et sp. n. is presently unknown.
Geographic distribution: this species is only known from the type locality: Mount Halcon, Baco municipality, Oriental Mindoro province, Mindoro Island, Philippines.
4 Discussion
4.1 Relationships and phylogenetic considerations
The major characters of D. eximius gen. et sp. n. can be summarized as follows: (a) neck region provided with a small median gular sclerite; (b) abdominal sternum IX of male represented by an undivided plate; (c) abdominal sternum VIII of female not keeled; (d) vertex cone-shaped, raised above the pronotum; (e) thoracic nota and abdominal terga I-IX of the male with hump-backed posteromedian swellings, absent in the female; (f) tegmina not developed, and hind wing reduced to a small yellow lateral lobe; (g) fore femur straight, neither compressed nor curved at base; (h) ventral femoral carinae of male armed with small spines, whilst are unarmed in the female; (i) tibia with the area apicalis consisting of a depressed membranous posterior lateral region, and a strongly sclerotized central apical region; (j) euplantulae consisting of smooth-type attachment pads; (k) pretarsal claws pectination developed in the form of minute denticulations; (l) boundary between the metanotum and the first abdominal tergum well differentiated; (m) cercus strongly elongated and gently incurved, lacking an apical spine in both sexes.
The first three characters listed above (a–c) are apomorphies shared with the other taxa of Aschiphasmatidae, and attest that D. eximius gen. et sp. n. belongs to this lineage. In fact, these features, along with the presence of the pro- and mesospina, the prothorax with the anapleurite separated from the coxopleurite by a membrane, the absence of the radial sector in the hind wing radius of winged taxa, and the closed internal micropylar plate of the egg, constitute the set of seven putative synapomorphies of the group [6].
According to Zompro [13] the head of Aschiphasmatidae is typically short, broad, and dorsoventrally flattened. In a few cases, for example in Aschiphasma Westwood, 1834, Ommatopseudes Günther, 1942 and Orthomeria Kirby, 1904, the vertex is moderately rounded and globular. The cone-shaped vertex represents therefore a new finding for this group, and can be considered a peculiar autapomorphy of the new taxon.
A sexually dimorphic character of D. eximius gen. et sp. n. is represented by the hump-backed swellings on the rear of the thoracic and abdominal terga. This feature is strongly developed in the male, whilst it is very indistinct in the female. Few other members of the group possess such seemingly apomorphic protrusions, notoriously the species of Dinophasma Uvarov, 1940 and Ommatopseudes Günther, 1942. It is interesting to note that across these taxa there is also a strong tendency towards the reduction and loss of the wings. In the specific case of D. eximius gen. et sp. n., only highly reduced hind wings are present. This condition is rare within Aschiphasmatidae, where can be observed also in Abrosoma apterum Redtenbacher, 1906. However, microptery is probably homoplasious in this phylogenetic context, and might not be a synapomorphy between these two species. In fact, the distinctive shape and coloration of the hind wings, particularly obvious in the male, represent autapomorphies of the new taxon, which are not known for any other representative of the group.
The structure of the fore femur has been used to investigate the intraphyletic relationships of the Aschiphasmatidae [3,13]. In most taxa the fore femur is compressed and incurved at the base, a condition considered among the groundplan apomorphies of the Euphasmatodea [6]. In Anoplobistus Bragg, 2001, the fore femur features only a slight curvature, whilst in Aschiphasma, Dajaca Brunner von Wattenwyl, 1893, Leurophasma Bi, 1995, and Orthomeria, it is neither significantly curved nor compressed at the base. With regard to the Euphasmatodea, the latter two character states can be interpreted as the result of a secondary modification. In fact, Bradler and Buckley [14] have correlated the non-incurved condition of the fore femora with secondarily ground-dwelling forms, and diurnal species characterized by aposematic colouration. D. eximius gen. et sp. n. features straight fore femora, in combination with a colour pattern which is strikingly rich in contrast, including typical aposematic colours like black and yellow. These features, along with the well-differentiated openings of the prothoracic exocrine glands, suggest that the new taxon might rely more heavily on its chemical defence system rather than on camouflage. Another interesting trait of the femora of D. eximius gen. et sp. n. concerns to the presence of sharp spikes. Bragg [3] noted that the ventral femoral carinae of most Aschiphasmatidae are equipped with some small spines. The situation found in the new taxon is somewhat atypical, since this spination is strongly developed in the male, but entirely missing in the female. This conspicuous difference between the sexes indicates that the function of these spines might be primarily related to aspects of the reproduction of the species, rather than defence against predators. Specifically, spines could be involved in aggressive interactions between males competing for access to females, and/or to achieve and maintain a specific position of the pair during copulation.
The tibial area apicalis is another groundplan characteristic of the Phasmatodea, and occurs approximately in one third of the extant species [6]. Tilgner [4] distinguishes between the membranous area apicalis of Timema Scudder, 1895, and the plate-like (or strongly sclerotized) area apicalis of the Euphasmatodea. However, Bradler [2] showed that members of the euphasmatodean genus Agathemera Stål, 1875 exhibit an intermediate condition: their area apicalis features both a membranous and a strongly sclerotized field. This latter model is here reported within Aschiphasmatidae. Other members of this lineage apparently present a fully sclerotized area apicalis [6,13]. However, as pointed out by Tilgner et al. [15], this character is still poorly documented among euphasmatodean taxa, and further morphological studies, especially with the use of the scanning electron microscope, would be highly desirable to evaluate the significance of these differences.
The fine-structural characteristics of the locomotory attachment devices of the Phasmatodea has been the subject of a recent study by Beutel and Gorb [16]. Groundplan features include a five-segmented tarsus equipped with well-developed attachment pads (arolium and euplantulae), whose flexible surfaces are densely covered with acanthae. Among the insects, this type of microstructure has been found also in the Mantophasmatodea, and is regarded as a possible synapomorphy of these two orders. In addition, unlike Timema, the adhesive surface of the arolium of the Euphasmatodea is smooth, apparently as a result of a secondary loss of the acanthae within this group [16]. Likewise, D. eximius gen. et sp. n. displays no acanthae in the arolium, even though its surface is not perfectly smooth. Most remarkably, acanthae are also absent in the euplantulae, which instead exhibit a smooth surface. This type of pad structure represents a groundplan characteristic of polyneopteran insects, which is found for example in the Dictyoptera, Grylloblattodea, and Orthoptera [17], and constitutes an unexpected finding for the Phasmatodea. It is possible that Aschiphasmatidae have secondarily lost the acanthae also in the euplantulae, but interestingly, the specialized honeycomb microstructure displayed by the new taxon is most similar to the condition observed in certain Orthoptera [18].
One of the major features that characterize the Aschiphasmatidae is the presence of distinctly multidentate-pectinate pretarsal claws. This characteristic is found uniformly within the lineage, with the notable exception of the members of Dajaca where the claws are simple [3], as reported in Timema and almost all other taxa of Euphasmatodea [4,6]. The character state expressed by Dajaca has been interpreted as a plesiomorphy, which would indicate a basal position of this genus within Aschiphasmatidae [6,13]. However, the highly reduced condition observed in D. eximius gen. et sp. n. brings into question the suggested ancestral placement of Dajaca. In fact, the minute denticulations of the new taxon might represent an intermediate state between the well-developed pectination and a secondary loss of the pectination. Therefore, the reduction and absence of pectination could be considered apomorphic characters for this group. The condition found in the new taxon has been also observed in other two enigmatic, closely related Aschiphasmatidae species: Dinophasma braggi (Zompro, 2004) from Borneo, and Dinophasma maalon Gottardo, 2007 from the Philippines. As pointed out by Gottardo [8], these two species differ significantly from the other members of Dinophasma Uvarov, 1940, which possess the typical well-developed pectination. The presence of reduced pectination could be a synapomorphy of D. eximius gen. et sp. n. and the two above-mentioned species.
Another important apomorphy shared by the Euphasmatodea is represented by the fusion of the metanotum with the first abdominal tergum [6,14,19]. As a consequence, the boundary that separates the two terga is typically structured as a faint suture, sometimes hard to distinguish. Conversely, in D. eximius gen. et sp. n. this boundary is well defined; particularly, the hind margin of the metanotum appears developed in the same manner to that of the adjacent terga. This feature might represent a plesiomorphic character state. In Timema, distinct membranous cuticle separates the concerned terga [15]. Once additional specimens of D. eximius gen. et sp. n. will be available for dissection, it will be possible to establish if membranes are also present between the metanotum and first abdominal tergum of this new taxon.
The cercus of D. eximius gen. et sp. n. is characterized by a cylindrical structure in both sexes, which is the most common condition among Aschiphasmatidae, including Dajaca. Only in a minority of the taxa, i.e. Anoplobistus, Aschiphasma, Eurybistus Bragg, 2001, and Orthomeria, males possess the more derived condition of distinctly flattened cerci. However, in both cases, the cercus of the male is usually incurved and equipped with a small spine or tooth in the apex, whilst that of the female is more straight, slightly smaller, and unarmed. These specializations of the male cercus are considered apomorphic with regard to the Aschiphasmatidae [6]. The male of D. eximius gen. et sp. n., which lacks the apical cercal spine, exhibits the plesiomorphic state. In addition to this feature, the cerci of the new taxon are peculiar for their considerable length in both sexes, which is a rare feature in this group of stick insects, and most likely apomorphic.
From the comparative analysis of the morphological characters, it appears that D. eximius gen. et sp. n. shares probable apomorphies (see characters e, g, and k) with the genera Dajaca, Dinophasma, and Ommatopseudes, that could suggest a close evolutionary relationship between these taxa.
4.2 Compendium of biodiversity information
A preliminary checklist of Philippine Aschiphasmatidae was recently drafted by Gottardo [8]. The present study increases the records to five genera and six species, expanding the geographical distribution of the group. The type locality of D. eximius gen. et sp. n., Mount Halcon on the eastern half of Mindoro island, represents the third highest mountain in the Philippines, but so far is known to harbour only few other phasmatodean taxa, specifically: four members of Lonchodinae: Lonchodiodes atrovirens Hennemann & Conle, 2007, Lonchodiodes eurycanthoides Hennemann & Conle, 2007, Manduria halconensis Hennemann & Conle, 1997, and Mithrenes mindorensis Hennemann & Conle, 2007; a representative of the Clitumninae, Phobaeticus philippinicus (Hennemann & Conle, 1997); one Obriminae, Trachyaretaon echinatus (Stål, 1877); and finally a Phylliinae, Phyllium (Phyllium) mindorense Hennemann, Conle, Gottardo & Bresseel, 2009 [11,20–22]. Most of these species appear to be Mindoro endemic, and future studies on this locality will certainly raise the number of taxa represented.
The enigmatic stick insect taxon described in this article also highlights the importance of the Philippines as an area featuring important endemic species of this polyneopteran insect order. The Philippine phasmatodean fauna, however, still remains poorly documented when compared to neighboring Asian areas such as Borneo or Peninsular Malaysia, whose faunas were recently revised [3,23]. Further studies are therefore urgently required, especially considering the exceptional loss of terrestrial ecosystems which is being experienced by the Philippines, representing the second ‘hottest hotspot’ of the biodiversity on Earth [24].
Disclosure of interest
The author declares that he has no conflicts of interest concerning this article.
Acknowledgements
The author wishes to express his gratitude to Sven Bradler for the helpful discussions on the morphological characters; to Oskar V. Conle, who has made available the material for study; and to Jerome Constant, who has translated the abstract from English to French.