

1 Introduction
It is commonly accepted by most authors that the discovery of the first element of the family Pseudochactidae by Gromov [1] represented one of the most remarkable scorpion discoveries of the 20th century [2–5]. Pseudochactas ovchinnikovi Gromov, 1998, was found in an isolated mountainous region of southeastern Uzbekistan and southwestern Tajikistan in Central Asia. This scorpion shares some features with both buthid and non-buthid scorpions, but it also displays a number of remarkable characters unique among extant scorpions.
Subsequent authors have not reached a consensus concerning the phylogenetic position of this enigmatic scorpion [2–5]; for details refer to Lourenço [6]. Another controversial aspect surrounding the pseudochactid lineage concerns its time and region of origin. Fet et al. [4] speculated about a possible Pangaean origin (Permian-Triassic), whereas Prendini et al. [5] estimate from the family's restricted geographical range and basal phylogenetic position that it should be considered as a palaeoendemic, a relict of an earlier scorpion fauna living in a more mesic environment. The subsequent discovery of new elements belonging to the pseudochactid lineage in Southeast Asian limestone caves totally re-opened this question.
Troglokhammouanus steineri Lourenço, 2007, was described from karst caves in Laos [6]. Subsequently, karst cave systems in Vietnam have been prospected for scorpions and several specimens of a new pseudochactid scorpion were collected [7,8]. A new genus, Vietbocap Lourenço & Pham, 2010, and two species, V. canhi Lourenço & Pham, 2010 and V. thienduongensis Lourenço & Pham, 2012, were described. These represented the first true troglobitic elements for the family Pseudochactidae. Very recently, two more specimens of Vietbocap were collected in the Tham Nam Lot cave, located in the Laotian region of the Khammouan karst. This represents a yet another new troglobitic species, but presents an even greater degree of adaptation to cave life. In account of the marked differences between Vietbocap and the other two genera of pseudochactids, a new subfamily, Vietbocapinae subfam. n., is proposed here to accommodate the species of this genus.
Four of the five known pseudochactid species, and two out of the three known genera are restricted to caves within the Khammouan-Ke Bang karst, located across the border between Laos and Vietnam (Fig. 1). This suggests that this region may represent an epicentre of diversity for the family Pseudochactidae. Moreover, this area probably played a role of a refuge or centre of endemism for elements of this family during past geological times. New phylogenetic or biogeographical considerations are not proposed here, since these aspects have already been discussed in detail by Lourenço [6]. Synthetic information on the orogeny and geodynamics of South East Asia can be found in Lourenço and Pham [7].
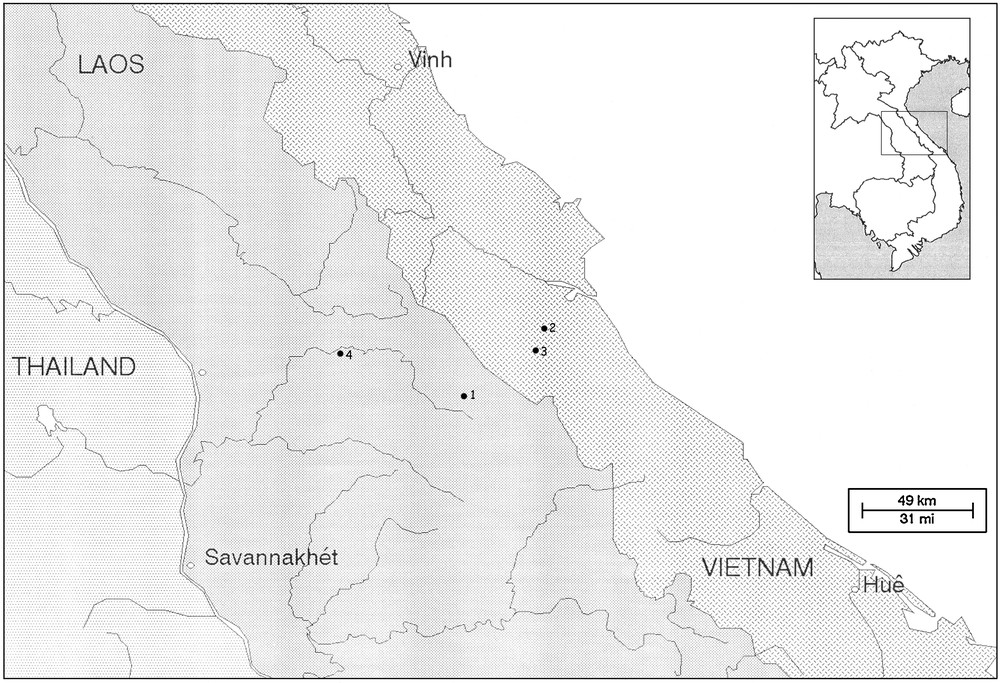
Map of the Khammouan karst system showing the location of the caves: 1. Tham Xe Bangfai cave. 2. Tien Son cave. 3. Thien Duong cave. 4. Tham Nam Lot cave.
2 Methods
Scorpions were collected in the caves with the help of standard electric frontal lights. They were found on cave walls and on the clay floor, approximately 350–400 m from the main cave entrance (Fig. 2). Measurements and illustrations were made using a Wild M5 stereomicroscope with a drawing tube and an ocular micrometer. Measurements follow those of Stahnke [9] and are given in mm. Trichobothrial notations are those developed by Soleglad and Fet [10] and the morphological terminology mostly follows that of Hjelle [11] and Lourenço [7,12].

Tham Nam Lot cave, interior view, showing Vietbocap lao sp. n. in its natural environment (photo L. Deharveng).
3 Annamite mountain range and Khammouan karst system
In Central Laos/Vietnam, the dominant geological feature is the Truong Son Range. This string of mountains and plateaus, also known as the Annamite Mountain Range, is roughly 1200 km long and 50–75 km wide, intersected by passes and lowlands. Most of its hills lie at elevations of 500–2000 m, and for much of its distance, they run parallel to the central coastline, straddling the border between Laos and Vietnam. The Truong Son Range is a transitional region between the subtropical communities of the North and the tropical ones of the South, and it harbours many endemic species [13,14]. It can be divided into three regions:
- (i) the Northern Truong Son, much of which is composed of ancient marine basins that have been uplifted and are now heavily eroded and form the characteristic sharp karst ridges and peaks, with extensive systems of caves, tunnels, underground rivers and streams;
- (ii) the Central Truong Son, dominated by the Kon Tum Massif-an enormous, largely granitic formation, which is among the oldest exposed formations in Southeast Asia;
- (iii) and the Southern Truong Son, including Vietnam's remaining uplands, with Dac Lac, Da Lat and Di Linh Plateaus-a series of eroded granite and basalt plateaus dotted with isolated peaks.
The Central Laos/Vietnam region comprises the most extensive tracts of limestone karst habitat in Asia after those of southern China. This unique karst system (290–255 My) was probably uplifted in the early Triassic and it differs substantially, in terms of both geology and habitat, from adjacent regions [15–17]. This is the oldest major karst area in Asia. It has been subject to massive tectonic changes and comprises a series of rock types that are interbedded in complex ways. Probably as many as seven different major levels of karst development have occurred as a result of tectonic uplift and changing sea levels. Thus the karst landscape is extremely complex with high geodiversity and many geomorphic features of considerable significance [17,18].
4 Taxonomic treatment
Family Pseudochactidae Gromov, 1998.
Subfamily Vietbocapinae subfam. n.
Diagnosis: Pair of circumocular sutures weak or vestigial; median and lateral ocelli absent; metasomal segment V with weakly marked ventrosubmedian carinae and lacking a ventromedian carina; fixed and movable fingers strongly curved; tibial spurs absent from all legs.
Type genus, Vietbocap Lourenço & Pham, 2010.
Genus Vietbocap Lourenço & Pham, 2010.
Vietbocap lao sp. n. (Figs. 2, 3).
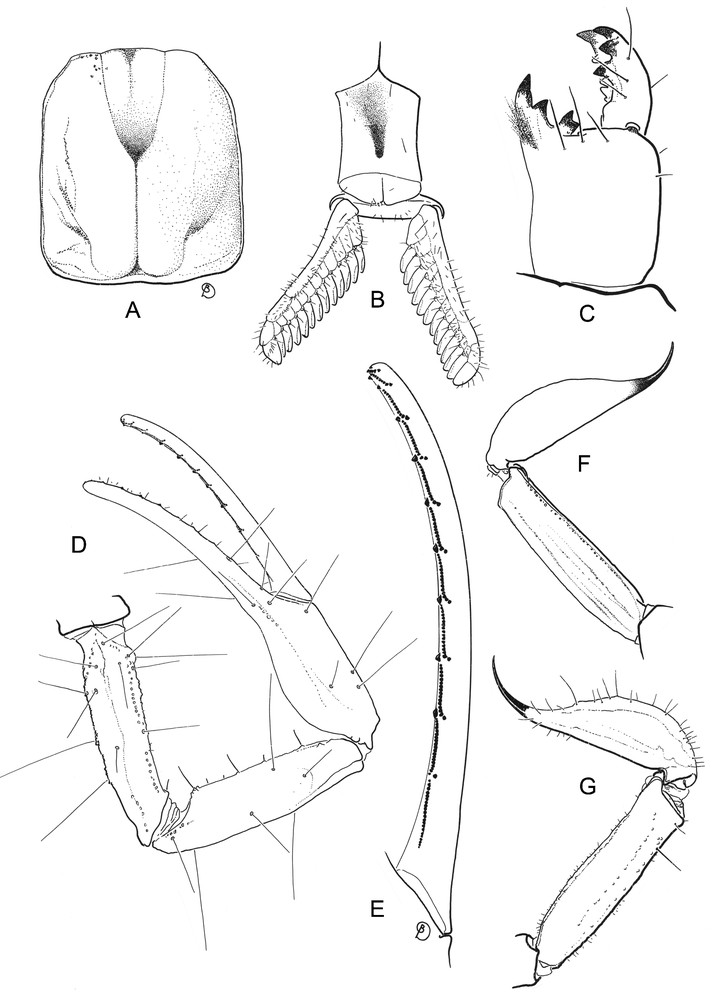
Vietbocap lao sp. n., female holotype. A. Carapace, dorsal aspect. B. Ventral aspect, showing sternum, genital operculum and pectines. C. Chelicera, dorsal aspect. D. Pedipalp, dorsal aspect, showing trichobothrial pattern. E. Movable finger of pedipalp chela with sub-rows of granules. F. Metasomal segment V and telson, lateral aspect. G. Idem for Vietbocap canhi, female holotype (from Lourenço and Pham [7]).
Diagnosis: anterior margin of carapace only slightly depressed, with a weak convexity. Lateral ocelli absent. Pair of circumocular sutures vestigial, with a broad U-shaped configuration; surface within the circumocular sutures less marked than in the other species of the genus. Median ocelli absent; median tubercle represented by a smooth, slightly depressed zone. Anterosubmedial carinae absent from zone delimited by circumocular sutures. Type D trichobothrial pattern [10,19] with 35 trichobothria per pedipalp: 12 on femur, of which 5 dorsal, 4 internal and 3 external (d1, d4, d5 and i4 extremely reduced); 10 on patella, of which 3 dorsal, 1 internal and 6 external (est extremely reduced); ventral surface without trichobothria; 13 trichobothria on chela, of which 5 on manus, 8 on fixed finger (ib2 extremely reduced); dorsal trichobothria of femur with ‘beta-like’ configuration. Sternum pentagonal, type 1 [20], strongly compressed horizontally, slightly longer than wide, external aspect not flat, with a concave region, posteromedian depression round. Telotarsi each with several spinular setae, which are not clearly arranged in rows. Metasomal segment V with a weakly marked pair of ventrosubmedian carinae; no ventromedian carina between ventrosubmedian carinae. Fixed and movable fingers strongly curved; dentate margins each with median denticle row comprising 10-11 oblique granular sub-rows; internal and external accessory granules at base of each sub-row. Pectines with 13-13 teeth. Respiratory spiracles small, semi-oval to round. Pro- and retrolateral pedal spurs present on legs I-IV. Tibial spurs absent from all legs.
Type material: female holotype, female paratype. Laos: Khammouane: Ban Naden: Tham Nam Lot cave (X = 105.385598; Y = 17.504969; alt = 176 m), 350–400 m from main cave entrance, 08/XI/2011 (L. Deharveng & A. Bedos) collected by hand (field number Lao11-46). Holotype and paratype will be deposited in the collections of the Muséum national d’Histoire naturelle, Paris.
Ecological notes: cave locally humid, composed of concretions and limestone; gallery of average size, (5–15 m length by 5–12 m high); no colony of bats observed; scorpions collected in zones sheltered from air currents, one found under a stone, the other moving over the concretions and limestone. The scorpions can run very rapidly. The habitat is largely mineral and no potential food sources for the scorpions were observed.
Etymology: the specific name refers to the native habitants of Laos.
Description based on female holotype and female paratype (measurements given after the description).
Colour: General coloration extremely pale yellow, almost white; much paler than the other two species of the genus; cheliceral teeth, telson tip and rows of granules on pedipalp fingers reddish.
Morphology: Chelicerae: dorsal edge of fixed finger with four denticles (basal, medial, subdistal, distal); ventral edge with four to five strongly reduced denticles; movable finger with three denticles (medial, subdistal, external distal) on dorsal edge, without basal denticles; ventral edge with four to five reduced denticles; external distal denticle slightly smaller than internal distal denticle; ventral aspect of fingers and manus with numerous macrosetae. Carapace: anterior margin only slightly depressed, with a weakly marked convexity; lateral ocelli absent; median ocular tubercle represented by a smooth and slightly depressed zone; median ocelli absent; interocular furrow obsolete. One pair of vestigial circumocular sutures with a broad U-shaped configuration. Anteromedian and posteromedian furrows shallow; posterolateral furrow shallow, weakly curved; posteromarginal furrow narrow, shallow. Carapace almost totally smooth, except for some isolated granules anteriorly and on postero-lateral edges. Pedipalp segments apilose. Femur with five discernible carinae, all weak; intercarinal surfaces smooth. Patella with six discernible carinae; ventrointernal carinae with one to two spinoid granules; intercarinal surfaces smooth. Chela with dorso-external carinae vestigial; ventral carina absent; tegument smooth. Fixed and movable fingers strongly curved; dentate margins each with median denticle row comprising 10–11 oblique granular sub-rows; each sub-row comprising several small granules and internal and external accessory granules. Trichobothria orthobothriotaxic, Type D [10,19], ‘beta-like’ configuration, d2 situated on dorsal surface, d3 and d4 in same axis of the femur, parallel and closer to dorsoexternal carina than is d1, angle formed by d1, d3 and d4 opening toward internal surface; totals: femur 12 (5 dorsal, 4 internal, 3 external); patella 10 (3 dorsal, 1 internal, 6 external); chela 13 (5 on manus, 8 on fixed finger); db slightly displaced over fixed finger. Legs I to IV: tibiae without spurs; basitarsi each with a pair of pro- and retrolateral spurs; telotarsi each with several spinular setae, not clearly arranged in rows. Sternum pentagonal, type 1 [20], strongly compressed horizontally, slightly longer than wide, external aspect not flat, with a concave region, posteromedian depression round. Pectines each with 3–4 distinct marginal lamellae and 7–8 well-delineated median lamellae in female. Fulcra absent or vestigial. Pectinal tooth count: 13–13 in holotype and paratype. Genital operculum divided longitudinally in two semi-oval plates. Mesosoma: pre-tergites smooth and shiny; post-tergites II-VI smooth, apart from some minute granules; VII with a few granules and a pair of dorso-submedian and dorsolateral carinae, reaching posterior edge of segment. Sternites almost entirely smooth, acarinate; surfaces with scattered macrosetae; distal margins with sparse row of macrosetae; respiratory spiracles small, semi-oval to round. Metasoma with a few short macrosetae. Ten carinae on segments I to III; intermediary carinae incomplete on III; eight carinae on segment IV; four on segment V. Dorso-submedian carinae moderately developed on segments I-IV, absent on segment V; spinoid granules absent. Other carinae moderately to weakly developed on segments I-V. Telson long and not bulbous; vesicle smooth on all faces; aculeus shorter than vesicle and weakly curved, without a subaculear tubercle ventrally. Form of venom glands unknown.
Measurements (in mm) of female holotype of Vietbocap lao sp. n. and female paratype of Vietbocap canhi. Total length 23.4/21.3. Carapace: length 3.2/2.8; anterior width 2.2/1.8; posterior width 3.1/2.9. Mesosoma length 4.9/6.4. Metasomal segments: I, length 1.4/1.0, width 1.4/1.2; II, length 1.5/1.2, width 1.3/1.0; III, length 1.6/1.4, width 1.3/0.9; IV, length 2.2/1.7, width 1.2/0.8; V, length 4.1/3.2, width 1.3/0.8, depth 1.1/0.8. Telson length 4.5/3.6; vesicle length 3.4/2.2, width 1.4/1.0, depth 1.2/0.9. Pedipalp: femur length 3.8/3.1, width 0.9/0.7; patella length 4.1/3.2, width 0.9/0.9; chela length 6.7/5.8, width 1.2/1.0, depth 1.1/0.9; movable finger length 4.4/3.9.
Geographic distribution: Only known from the type locality.
5 Relationships
Although similar in general morphology, Vietbocap lao sp. n. can be distinguished from both V. canhi and V. thienduongensis by a number of features:
- (i) several distinct morphometric values, see measurements;
- (ii) circumocular sutures less marked, almost vestigial;
- (iii) chela fingers with 10-11 sub-rows of granules vs. 7 in V. canhi and 8 in V. thienduongensis;
- (iv) pedipalps less carinated and granulated;
- (v) pectines more elongated, with 13 teeth (presumably 16 to 17 in males) vs. 7 in V. canhi.
6 Discussion
The three known species of Vietbocap all present a very similar general morphology and probably derive from a common ancestral lineage which colonized the caves of the Khammouan-Ke Bang karst system. Two possible scenarios can be suggested. A common epigean ancestor colonized the different caves independently, supposing that when this happens the caves were already isolated. Alternatively, one or several closely related epigean ancestors colonized the entire Khammouan karst system, which is the oldest major karst area in Asia (290–255 My) and has been subject to massive tectonic changes since the early Triassic [15–17]. Previously connected caves subsequently became isolated, leading to speciation between different Vietbocap populations. However, our present knowledge of the scorpion fauna of the caves in the Khammouan karst system is extremely preliminary. Besides, nothing is known about possible epigean or endogean elements of Pseudochactidae in the regions close to the karst system.
The species of Vietbocap present a very marked degree of adaptation to cave life, particularly so in Vietbocap lao sp. n. Their situation is comparable to that of the cave elements of the genus Typhlochactas Mitchell, 1971 (family Typhlochactidae), distributed in Mexican caves. In the case of Typhlochactas, however, at least three litter species have been described, besides the known caves species [21]. All the pseudochactid species discovered in the Khammouan-Ke Bang karst system were found at very significant distances from the main cave entrance: 200 m for V. canhi, 350–400 for V. lao sp. n. and 1800 m for V. thienduongensis. Unpublished recent data for the latter confirms distances of up to 2100 m, which are the longest distances from a cave entrance known for any scorpion.
T. steineri was also found at a remarkable distance (2000–3000 m) from the main cave entrance, but its morphological characteristics are not those of a troglobitic element. Besides, collections at the same site also included several insects, spiders, harvestmen, isopods and millipedes. This suggests that other secondary entrances may exist in Tham Xe Bangfai cave, where T. steineri was found [6].
Disclosure of interest
The author declares that he has no conflicts of interest concerning this article..
Acknowledgements
I am most grateful to Louis Deharveng and Anne Bedos (CNRS/MNHN) for collecting the material used in the description of the new species and for information on the ecology of the cave; Bernard Duhem (MNHN) for preparing the illustrations; Elise-Anne Leguin (MNHN) for the preparation of the plates; and to Mark Judson (MNHN) for corrections to the manuscript. Thanks are also extended to the ATM/MNHN for financial support; J. P. Hugot who organized the field work; the Nam Theun 2 Power Company (Vientiane, Lao PDR) for its logistical help in the field; Dr B. Bouahom, Director of the National, Agricultural and Research Institute (NAFRI) for his interest in our investigation.
Appendix A Appendix A
Key to the known genera and species of Pseudochactidae.