

1 Introduction
The family Pseudochactidae Gromov, 1998 contains some of the most remarkable scorpions described in recent years. The first species to be discovered was Pseudochactas ovchinnikovi Gromov, 1998, found in an isolated mountainous region of southeastern Uzbekistan and southwestern Tajikistan, in Central Asia [1]. A second genus and species, Troglokhammouanus steineri Lourenço, 2007, was described from karst caves in Laos [2]. Although this species was found inside a cave, its morphological characteristics do not correspond to a troglobitic element. This Laotian species reopened the question about the origins and affinities of the Pseudochactidae and led to new biogeographical interpretations [2].
Very recently, scorpions have been prospected in karst cave systems in Vietnam, and several specimens of a new pseudochactid scorpion were collected in the Tien Son cave, which belongs to the Phong Nha system. These were described as a new genus and species, Vietbocap canhi Lourenço & Pham, 2010, which represents a true troglobitic element [3]. In recent months, new surveys in the cave systems of Vietnam have been carried out and again, another pseudochactid scorpion was collected in the Thien Duong cave, which belongs to the Vom cave system. The new species also belongs to the genus Vietbocap, and shows features of a true troglobitic element. The fact that three pseudochactid elements originating from caves within the same karst system have already been found in Laos and Vietnam, suggest that this region of Southeast Asia may represent a refuge or centre of endemism for this family.
New phylogenetic or biogeographical considerations are not proposed here, since these aspects have already been largely discussed by Lourenço [2]. More detailed information on the orogeny and geodynamics of South East Asia, and on the location, ecology and climate of the national park and caves, can be found in Lourenço & Pham [3].
2 Tien Son cave in the Phong Nha cave system and Thien Duong cave in the Vom cave system
Tien Son cave, where Vietbocap canhi was found is located in Son Trach Commune, Bố Trạch District (Fig. 1). The entrance is located 1 km from Phong Nha cave, at an altitude of 200 m. Tien Son cave is 980 m in length. A 10 m deep hole is situated 400 m from the entrance, after which a 500 m long underground cave is open solely to professional scientists. According to British speleologists, Tien Son Cave was created tens of millions years ago, when a water current holed this limestone mountain in Ke Bang. Following a series of rock movement, this mass was levered or lowered, blocking the current and creating what is now Tien Son Cave, while the flow of the underground river was redirected to Phong Nha Cave. Although Phong Nha and Tien Son Caves are located next to each other, there are no passages linking them [4].
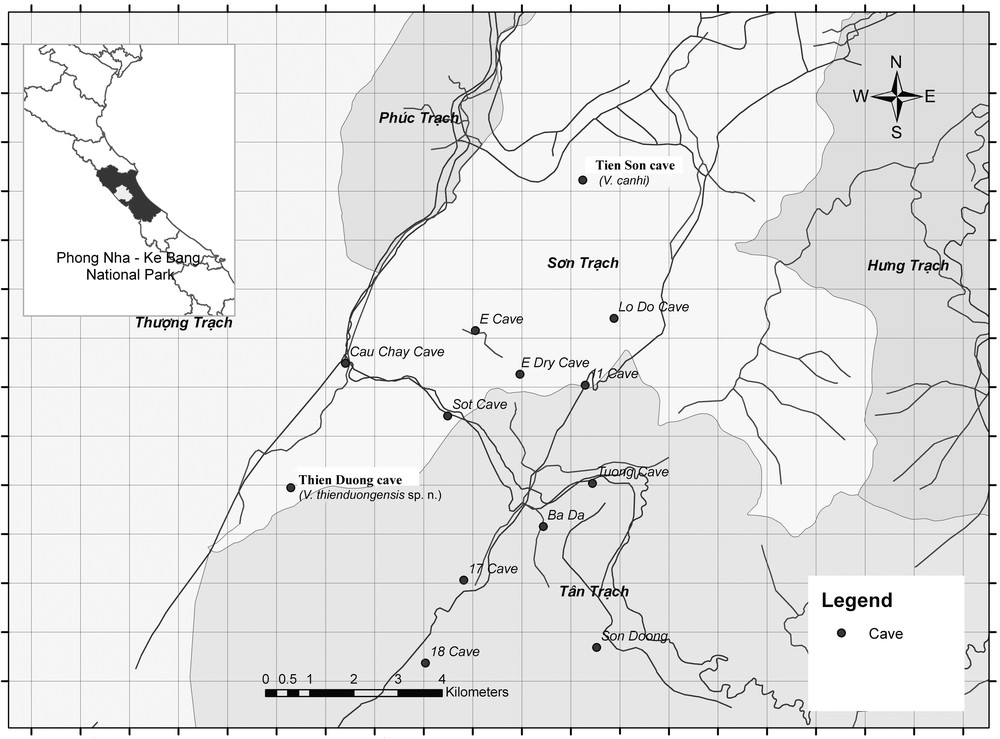
Map of the Phong Nha - Ke Bang National Park showing the locations of Tien Son and Thien Duong caves.
Thien Duong cave (Paradise cave), where the new species was found, is situated in Phong Nha-Ke Bang National Park, 60 km northwest of Dông Hói city (Fig. 1). Thien Duong cave is at an elevation of 200 meters above sea level, near the west branch of Ho Chi Minh Highway, in Son Trach Commune, Bo Trach District, Quang Binh Province, Vietnam. The cave was discovered by a local inhabitant in 2005 and initially the first 5 km of this cave were explored by scientists from the British Cave Research Association in 2005. More recently the whole extension of the cave was explored by the same Association. The cave is 31 km long, and in parts can reach 100 meters in height and 150 meters in width. There are two cave systems in Phong Nha-Ke Bang region: Phong Nha cave system and Vom cave system. However, these systems are totally isolated, with no geological connections being known between them [5].
The Phong Nha-Ke Bang karst is the oldest major karst area in Asia. It has been subjected to massive tectonic changes and comprises a series of rock types that are interbedded in complex ways. Probably as many as seven major levels of karst development have occurred as a result of tectonic uplift and changing sea levels, thus the karst landscape of PNKB is extremely complex with high geodiversity and many geomorphic features of considerable significance [3,5].
3 Methods
Scorpions were collected by scientists of the IEBR and the Phong Nha-Ke Bang National Park, while exploring the caves with the help of standard electric torches. They were found on the cave walls, approximately 1800 m from the main cave entrance (while V. canhi was found, under rocks, 200 m from the main entrance of Tien Son Cave). This is a new distance record from a cave entrance for a scorpion (Fig. 2). Measurements and illustrations were made using a Wild M5 stereomicroscope with a drawing tube and an ocular micrometer. Measurements follow those of Stahnke [6] and are given in mm. Trichobothrial notations are those developed by Soleglad and Fet [7] and the morphological terminology mostly follows that of Hjelle [8] and Lourenço [2,9].

Thien Duong cave, interior view, showing the walls where the new species was found.
4 Taxonomic treatment
Family Pseudochactidae Gromov, 1998
Genus Vietbocap Lourenço & Pham, 2010
Vietbocap thienduongensis sp. n. (Fig. 3)
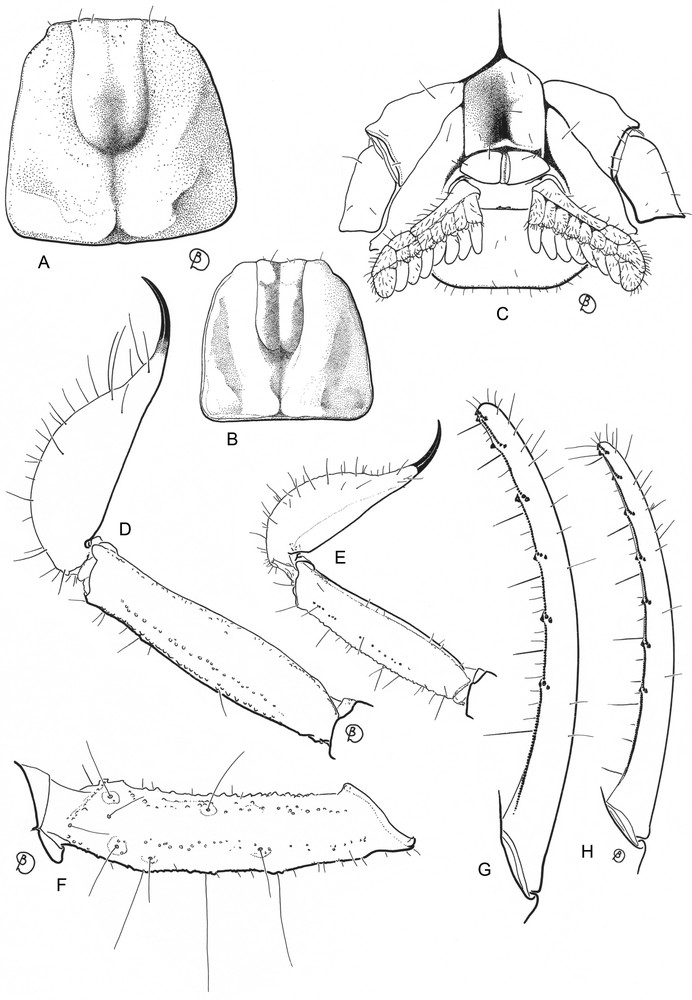
A, C, D, F, G. Vietbocap thienduongensis sp. n., male holotype. B, E, H. Vietbocap canhi, male holotype (from Lourenço & Pham [3]. A–B. Carapace, dorsal aspect. C. Ventral aspect, showing sternum, genital operculum, pectines and sternite III. D–E. Metasomal segment V and telson, lateral aspect. F. Femur, dorsal aspect, showing trichobothrial pattern. G–H. Movable finger of pedipalp chela with sub-rows of granules.
4.1 Diagnosis
Anterior margin of carapace only slightly depressed, with a weak concavity. Lateral ocelli absent. Pair of circumocular sutures complete in the posterior region to median ocular tubercle with a broad U-shaped configuration. Median ocelli absent; median tubercle represented by a smooth but not depressed zone. Anterosubmedial carinae absent from zone delimited by circumocular sutures. Type D trichobothrial pattern [7,10] with 35 trichobothria per pedipalp: 12 on femur, of which 5 dorsal, 4 internal and 3 external (d1, d4, d5 and i4 extremely reduced); 10 on patella, of which 3 dorsal, 1 internal and 6 external (est extremely reduced); ventral surface without trichobothria; 13 trichobothria on chela, of which 5 on manus, 8 on fixed finger (ib2 extremely reduced); dorsal trichobothria of femur with ‘beta-like’ configuration. Sternum pentagonal, type 1 [11], strongly compressed horizontally, slightly longer than wide, external aspect not flat, with a concave region, posteromedian depression round. Telotarsi each with several spinular setae, not clearly arranged in rows. Metasomal segment V with a weakly marked pair of ventrosubmedian carinae; no ventromedian carina between ventrosubmedian carinae. Fixed and movable fingers strongly curved; dentate margins each with median denticle row comprising seven oblique granular sub-rows; internal and external accessory granules at base of each sub-row. Respiratory spiracles small, semi-oval to round. Pro- and retrolateral pedal spurs present on legs I-IV. Tibial spurs absent from all legs.
4.2 Type material
Male holotype, male paratype. Vietnam, Quang Binh Province, Phong Nha - Ke Bang National Park, Thien Duong cave (106° 22′E–17° 52′N), mid section of cave (1800 m from cave entrance), 9/VIII/2011 (N.-K. Dang). Holotype deposited in the Muséum national d’Histoire naturelle, Paris. Paratype deposited in the Institute of Ecology and Biological Resources, Vietnam Academy of Science and Technology, Hanoi.
4.3 Etymology
Specific name refers to Thien Duong cave, where the new species was found.
Description based on male holotype and male paratype (measurements given after the description).
4.4 Colour
General coloration yellow, less pale than V. canhi; cheliceral teeth, telson tip and rows of granules on pedipalp fingers dark reddish.
4.5 Morphology
Chelicerae: dorsal edge of fixed finger with four denticles (basal, medial, subdistal, distal); ventral edge with 3–4 very reduced denticles; movable finger with three denticles (medial, subdistal, external distal) on dorsal edge, without basal denticles; ventral edge with 4–5 reduced denticles and a very weak serrula; external distal denticle smaller than internal distal denticle; ventral aspect of fingers and manus with numerous macrosetae. Carapace: anterior margin only slightly depressed with a weakly marked concavity; lateral ocelli absent; median ocular tubercle represented by a smooth and not depressed zone; median ocelli absent; interocular furrow obsolete. One pair of weakly marked circumocular sutures with a broad U-shaped configuration, also complete behind median ocular tubercle. Anteromedian and posteromedian furrows shallow; posterolateral furrow shallow, weakly curved; posteromarginal furrow narrow, shallow. Carapace almost totally smooth, except for some isolated granules anteriorly. Pedipalp segments apilose. Femur with five discernible carinae, all weak; intercarinal surfaces smooth. Patella with six discernible carinae; ventrointernal carinae with some spinoid granules; intercarinal surfaces smooth. Chela with dorso-external and ventral carinae weakly marked; tegument smooth. Fixed and movable fingers strongly curved; dentate margins, each with median denticle row comprising seven oblique granular sub-rows; each sub-row comprising several small granules and internal and external accessory granules. Trichobothria orthobothriotaxic, Type D [7,10], ‘beta-like’ configuration, d2 situated on dorsal surface, d3 and d4 in same axis of the femur, parallel and closer to dorso-external carina than is d1, angle formed by d1, d3 and d4 opening toward internal surface; totals: femur, 12 (5 dorsal, 4 internal, 3 external); patella, 10 (3 dorsal, 1 internal, 6 external); chela, 13 (5 on manus, 8 on fixed finger). Legs I to IV: tibiae without spurs; basitarsi each with a pair of pro- and retrolateral spurs; telotarsi each with several spinular setae, not clearly arranged in rows. Sternum pentagonal, type 1 [11], strongly compressed horizontally, slightly longer than wide, external aspect not flat, with a concave region, posteromedian depression round. Pectines each with 3–4 distinct marginal lamellae and 7–8 well-delineated median lamellae in male. Fulcra absent or vestigial. Pectinal tooth count: 8–8 in males. Genital operculum completely divided longitudinally; genital plugs observed in male. Mesosoma: pre-tergites smooth and shiny; post-tergites II–VI smooth, apart from some minute granules; VII with a few granules and a pair of dorso-submedian and dorsolateral carinae, reaching posterior edge of segment. Sternites almost entirely smooth, acarinate; surfaces with scattered macrosetae; distal margins with sparse row of macrosetae; respiratory spiracles small, semi-oval to round. Metasoma with a few short macrosetae. Ten carinae on segments I to III; eight carinae on segment IV; four on segment V. Dorso-submedian carinae moderately developed on segments I–IV, absent on segment V; spinoid granules absent. Other carinae moderately to weakly developed on segments I–V. Telson long and slightly bulbous; vesicle smooth on all faces; aculeus shorter than vesicle and weakly curved, without a subaculear tubercle ventrally. Form of venom glands unknown. Geographic distribution: only known from the type locality.
4.6 Measurements (in mm) of male holotype of Vietbocap canhi and male holotype of Vietbocap thienduongensis sp. n.
Total length 22.4/27.3. Carapace: length 2.9/3.6; anterior width 2.0/2.2; posterior width 3.2/3.5. Mesosoma length 5.5/6.7. Metasomal segments: I, length 1.2/1.4, width 1.4/1.6; II, length 1.4/1.7, width 1.3/1.4; III, length 1.5/2.0, width 1.2/1.4; IV, length 2.1/2.3, width 1.1/1.3; V, length 3.9/4.8, width 1.1/1.3, depth 0.9/1.2. Telson length 3.9/4.8; vesicle length 2.4/3.6, width 1.3/1.6, depth 1.2/1.4. Pedipalp: femur length 3.8/4.5, width 0.9/0.9; patella length 3.6/4.3, width 1.1/1.2; chela length 7.1/8.3, width 1.2/1.3, depth 1.0/1.2; movable finger length 4.2/4.6.
5 Relationships
Vietbocap canhi and Vietbocap thienduongensis sp. n. are rather similar in morphology. However, the new species can be distinguished from V. canhi by a number of features: (i) bigger size and distinct morphometric values; (ii) complete and more strongly marked circumocular sutures; (iii) chela fingers proportionally shorter (ratios of chela length/movable finger length 7.1/4.2 = 1.69 for V. canhi and 8.3/4.6 = 1.80 for V. thienduongensis sp. n.) and with 8 sub-rows of granules vs. 7 sub-rows; (iv) sternum only slightly longer than wide (ratio 1.15 for V. canhi and 1.30 for V. thienduongensis sp. n.); (v) metasomal segments less carinated and granulated and with a weaker chaetotaxy; (vi) pedipalp carinae better marked; (vii) pectines shorter and more bulkier with 8 teeth.
Moreover, the caves where the species have been found are totally isolated from one other and belong to distinct cave systems. Given that the two caves are only a few kilometres apart, it can be suggested that both Vietbocap species may have a common epigean ancestor that colonized the caves independently.
6 Key to the known genera and species of Pseudochactidae
This is shown in Fig. 4.

Key to the known genera and species of Pseudochactidae.
Disclosure of interest
The authors declare that they have no conflicts of interest concerning this article.
Acknowledgements
We are most grateful to Bernard Duhem (MNHN, Paris) for preparing the illustrations; to Elise-Anne Leguin (MNHN, Paris) for the preparation of the plates; and to Mark Judson (MNHN, Paris) for corrections to the manuscript. The second author wishes to thanks Dang Ngoc Kien, Phung Thi Hong Luong, Dang Van An, Le Quang Tuan, and Ho Khanh for their help during the field survey; the Nature Conservation and Sustainable Natural Resource Management of the Phong Nha–Ke Bang National Park Region Project for logistic support during field studies and funds from the Vietnam-Germany Development Cooperation.