

1 Introduction
Ocean sunfish (Mola mola (L., 1758), Molidae, Tetraodontiformes) and angler (Lophius piscatorius (L., 1758), Lophiidae, Lophiiformes) are acanthomorph fishes differing considerably in their ecology and their external anatomy (Fig. 1). The first is one of the heaviest marine vertebrates (up to 2.3 tonnes [1]), a disc-shaped, laterally flattened pelagic fish able to dive several times each day from the surface to a depth of more than 100 m [1–4] while the second is a smaller, ventro-dorsally flattened bottom dweller, living at depths of 30–500 m [5–7]. Ocean sunfish are rare and cosmopolitan, while anglers are currently fished commercially by European fisheries. The skeletal anatomy of the species of these groups has been well described [8–10], but the characteristics of their internal organs are little studied. In classic works, organs were often removed from the abdominal cavity and studied separately [11–16]. Although a few similarities were noted, for example in the organization of the spinal cord which does not extend into the spinal canal [16–18], they were regarded as very different animals [19–23]. Until recently, angler and ocean sunfish were considered as unrelated species in hypotheses of teleostean interrelationships [19–23]. However, numerous studies on both DNA sequences (mitochondrial and nuclear genes) of Lophiiformes and Tetraodontiformes species showed that they were closely related [24–33] and share a common ancestor with Scatophagidae, Cepolidae and Priacanthidae [34,35]. Two other works [36,37] examined the problem of the phylogenetic position of these groups. Unfortunately, one of these works [36] did not include both Lophiiformes and Tetraodontiformes and thus did not challenge or corroborate a close relationship between them. In the other work [37], Lophiiformes and Tetraodontiformes do not appear to be closely related on the basis of five gene regions (nuclear and mitochondrial), but this result is quite unique, while ten different works [24–33] conducted by different teams on various genes (17 nuclear genes and 34 mitochondrial genes) corroborate a close relationship between Lophiiformes and Tetraodontiformes. If angler is a derived lophid species [7], but occupies a basal position within the Lophiiformes [38,39], ocean sunfish is a derived tetraodontiform fish [23,28,40]. Ideally, the internal anatomy of these species should also be compared to one of more basal tetraodontiforms, such as triggerfishes (Balistidae). Unfortunately, we cannot obtain large fresh specimens of this family to conduct the comparisons in similar conditions. We took the opportunity of having access to fresh ocean sunfish to conduct CT imaging and magnetic resonance imaging (MRI) investigations and to compare the internal anatomy of the ocean sunfish and the angler.

External morphology (left lateral views) of (A) an ocean sunfish (Mola mola, Molidae) and (B) an angler (Lophius piscatorius, Lophiidae). Scale bars indicate 20 cm.
Drawings from F. Dejouannet.
2 Materials and methods
2.1 Specimens studied
Four angler specimens ((an isolated head (Total Length) TL = 25 cm) and three whole specimens (specimen 1: TL = 32.5 cm, (Standard Length) SL = 27.6 cm; specimen 2: TL = 33 cm, SL = 28.5 cm and specimen 3: TL = 62 cm, SL = 57.3 cm)) were fished in the eastern Atlantic Ocean and purchased at the fish market auction of Concarneau (France). An ocean sunfish (TL = 142 cm, body depth without median fins = 175 cm, with the right eye punctured) was fished in the western part of the English Channel by local fishermen from Granville (50, France). It was collected by the show aquarium of Granville and then sent to the Laboratoire d’Anatomie Comparée, ONIRIS (Nantes, France) where it has been cast for the Muséum d’Histoire Naturelle of Nantes (a few plastiline remains are visible on Fig. 3B). The animal was conserved frozen in the Laboratoire d’Anatomie Comparée, ONIRIS. Median fins were cut to reduce the size of the specimen for the MRI and CT examinations. The MRI examinations were conducted in the Unité d’Imagerie Médicale, ONIRIS, the CT imaging was performed by Image-Et (Mordelles, France). The skeleton of this ocean sunfish will be prepared for exhibitions at the Muséum d’Histoire Naturelle of Nantes (France).

A. Sagittal image (CT) of ocean sunfish. B. 3-D Volume Rendering image (CT) of ocean sunfish, lateral view. an pt: anal pterygiophore; cc: cranial cavity; ec: eye cavity; g: gills; hms: haemal spine; in: intestine; j: jaws; li: liver; m: mouth; plr: plastiline remains; pre: punctured right eye (filled with plastiline); rpf: right pectoral fin; vc: vertebral column. Scale bars indicate 15 cm.
2.2 CT imaging
CT examinations were performed on a 16-slice Computed Tomography manufactured by Siemens (Sensation 16, dedicated to the veterinary environment and industry by Image-Et (Mordelles, France)). Helical acquisitions were performed with a collimation High Resolution (HR, 16 × 0.75 mm) after the Scout View. The constants were 100 kV and 180 mAs eff. Two primary reconstructions, 0.75 mm with a reconstruction increment of 0.4 mm, were made in filter B (Body): one in soft (B20), the other medium-hard (B60), both in the bone window (Window Center 450 HU, Window Width 1500 HU). The Field of View (FOV) of the reconstructions was 204 mm. Radial post-processing views were carried out from the primary reconstruction using a soft filter (B20) to obtain images in 3D Volume Rendering.
2.3 Magnetic resonance imaging
MRI examinations were also performed on each specimen. Here we used a 1 Tesla supraconductor magnet (Harmony Siemens). A standard body coil was employed along with a three planes localizer. The T1 (TR = 516–656 ms and TE = 13 ms) and T2 (TR = 3840–5170 ms and TE = 91–115 ms) sequences in three planes (virtual sagittal, horizontal (= coronal) and transverse) extending from the anterior extremity of the snout to the distal tip of the caudal fin were acquired after the specimens were thawed. The thickness of the slices (576 for M. mola) varied from 3–10 mm depending on our interest in the details of the sequence: for transverse sections (205 for M. mola, 210 for L. piscatorius), TR = 4020, TE = 102, FOV = 320 × 500, thickness of slices = 10 mm, for coronal sections (208 for M. mola, 285 for L. piscatorius), TR = 4020, TE = 102, FOV = 320 × 500, thickness of slices = 10 mm, for sagittal sections (271 for M. mola, 180 for L. piscatorius), TR = 5170, TE = 91, FOV = 375 × 500, thickness of slices = 10 mm. No contrast agent was used. Three-dimensional (3-D) MRI datasets were tested on images from ONIRIS (Nantes) and from the Digital Fish Library (DFL), http://www.digitalfishlibrary.org, (images from the DFL have been kindly provided by R. Berquist). Unfortunately, the resolution of images was not equivalent (Fig. 2) and 3-D reconstructions were not yet anatomically completely suitable. We are now working on 3-D datasets and hope being able to provide higher quality reconstructions in a near future.

Comparison of coronal images (MRI) of ocean sunfish. A. Image present on the DFL. B. image obtained in ONIRIS (Nantes). Scale bars indicate 7 cm.
3 Results
CT imaging and MRI are important techniques to investigate non-invasively the gross anatomy of specimens. Both provide information about internal structures which corroborates and completes previous observations [15–18,41] and MRI examinations of ocean sunfish and related species available in the DFL. In the present study, CT imaging has provided data about the mineralized structures of the angler while MRI has provided information on the soft anatomy of the ocean sunfish. Both have been complementary in documenting the organization of organs (Figs. 3–9). CT imaging has been essential here to determine the 3D the disposition of skeletal elements and to reveal their structure. Nevertheless, the cartilaginous skeleton of ocean sunfish [1,15] is poorly resolved by CT imaging (Fig. 3), but virtual sagittal, horizontal and transversal sections obtained by MRI delivered more detail than CT imaging: the brain, the gallbladder and the organization of the heart are more visible in MRI images (Figs. 5 to 9).
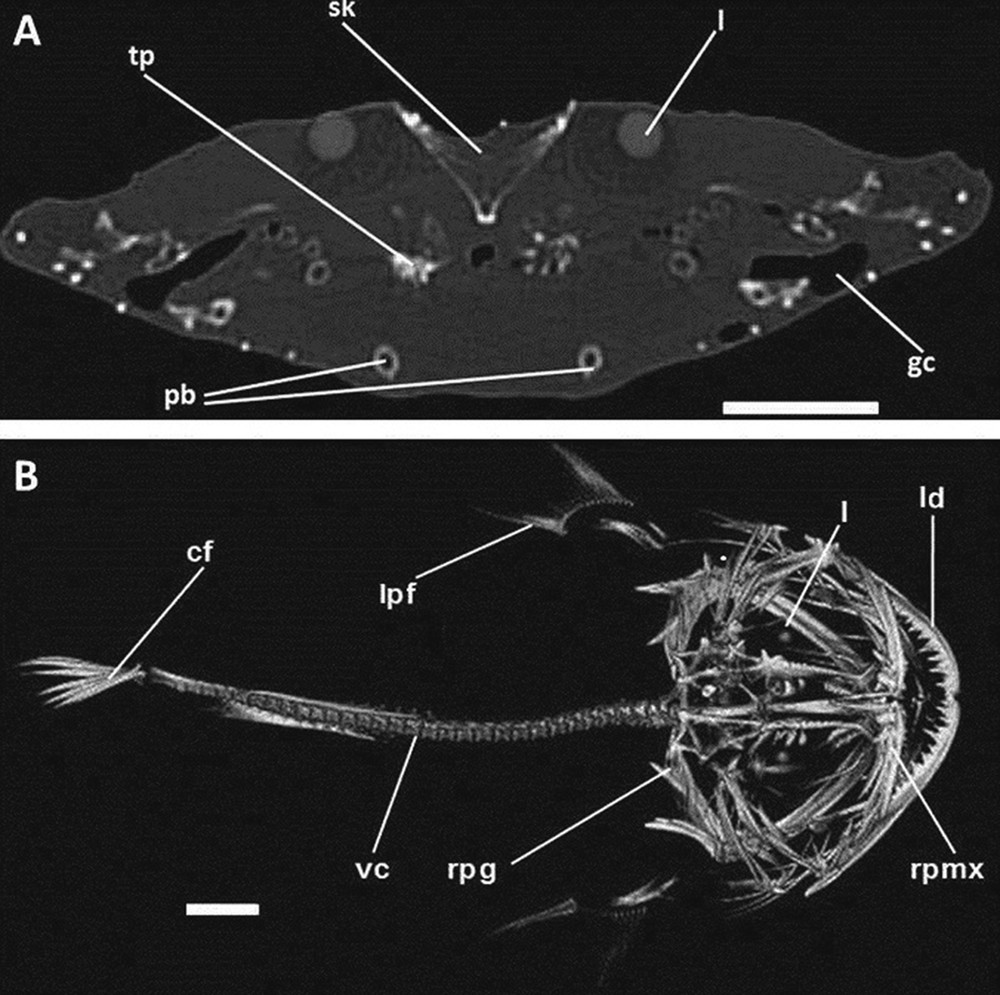
A. Transversal image (CT) of angler. B. 3-D Volume Rendering image (CT) of angler, dorsal view. cf: caudal fin; gc: gill chamber; l: lens of the eye; ld: left dentary; lpf: left pectoral fin; pb: pelvic bones; rpg: right pectoral girdle; rpmx: right premaxilla; sk: skull; tp: toothed plate; vc: vertebral column. Scale bars indicate 5 cm.

Sagittal image (MRI) of ocean sunfish. T2-weighted image. a: atrium; ba: bulbus arteriosus; br: brain; cc: cranial cavity; e: esophagus; g: gills; in: intestine; li: liver; pc: pericardiac cavity; sk: skull; st: stomach; th: thyroid; v: ventricle; va: ventral aorta. Scale bar indicates 7 cm.
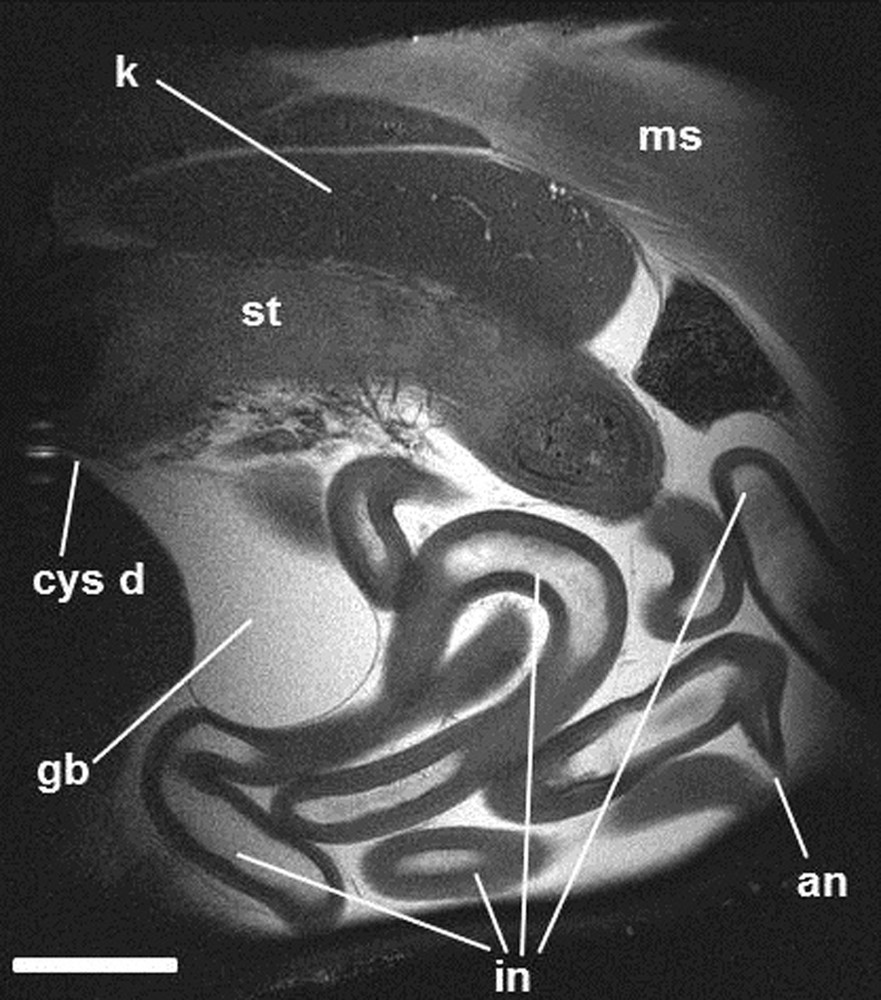
Sagittal image (MRI) of ocean sunfish.T2-weighted image. an: anus; cys d: cystic duct; gb: gallbladder; in: intestine; k: kidney; ms: muscles; st: stomach. Scale bar indicates 7 cm.
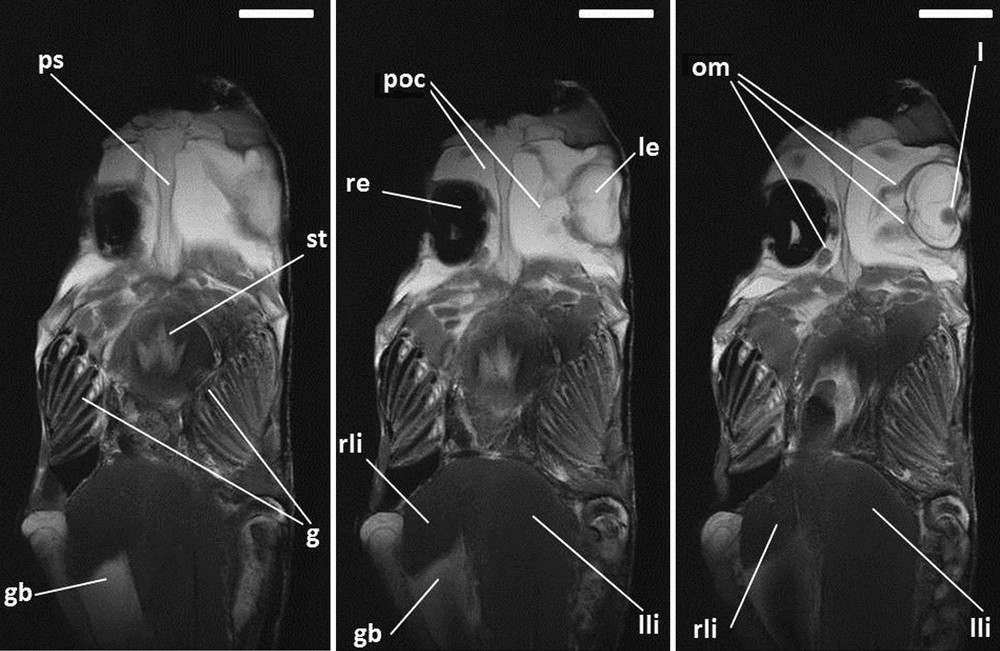
Successive horizontal images (MRI) of ocean sunfish. T2-weighted images. g: gills; gb: gallbladder; l: lens of the left eye; le: left eye; lli: left lobe of the liver; om: ocular muscles; poc: periocular cavities; ps: parasphenoid; re: right eye (punctured); rli: right lobe of the liver; st: stomach. Scale bars indicate 7 cm.

Successive horizontal images (MRI) of angler. T2-weighted images. a: atrium; ba: bulbus arteriosus; ca: pyloric caecum; gb: gallbladder; h: heart; li: liver; lin: large intestine; lli: left lobe of the liver; ms: muscles; sin: small intestine; st: stomach; v: ventricle; vc: vertebral column. Scale bars indicate 4 cm.
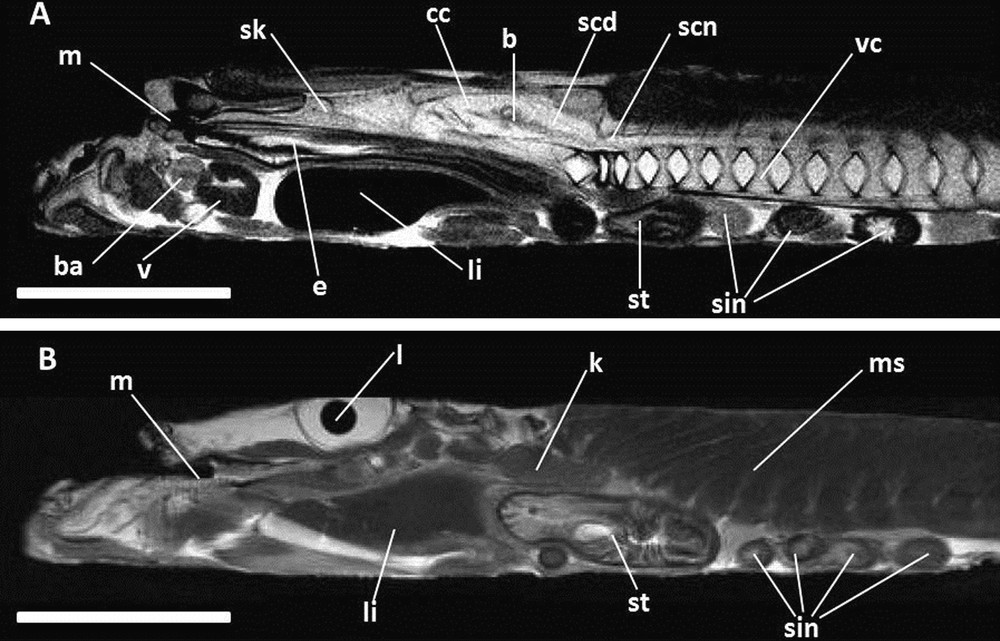
A. and B. Successive sagittal images (MRI) of angler.T2-weighted images. b: brain; ba: bulbus arteriosus; cc: cranial cavity; e: esophagus; k: kidney; l: lens; li: liver; m: mouth; ms: muscles; scd: spinal cord; scn: spinal canal; sin: small intestine; sk: skull; st: stomach; v: ventricle; vc: vertebral column. Scale bars indicate 5 cm.
In ocean sunfish, the brain occupies a small part of the cranial cavity (Fig. 5); it is surrounded by an abundant tissue (appearing in white in T2 weighting) (Fig. 5). The right eye, which is punctured in the specimen, is full of plastiline (Figs. 2 and 3) and appears in black with MRI (Figs 2 and 7). The heart is voluminous, with an enormous dorsally located atrium and an expanded pericardial cavity full of liquid (Fig. 5). This presence of liquid within the body of ocean sunfish is unusual and is clearly showed with MRI examination with T2 weighting, where liquids appear in white. Such pouches are present around each eye, which is surrounded by a large periocular cavity (Fig. 7) and around the coils of the intestine of ocean sunfish (Figs. 5 and 6). A large pouch full of liquid is also present along the ventral aorta antero-dorsally to the ventricle (Fig. 5); it may be the thyroid gland embedded in a blood lacuna. The swimbladder is absent and the stomach is distinct but not well developed (Figs. 5 and 6). A very long and coiled intestine occupies the main portion of the abdominal cavity (Figs. 3, 5 and 6). This organ coils in vertical planes, and no pyloric caecum is present. The liver is a large organ (Figs. 3, 5 and 7), composed of two lobes, with the left one being much more developed than the right one (Fig. 7). The gallbladder appears as a developed and rounded vesicle (Fig. 6) present on the right side between the two hepatic lobes (Fig. 7). The opening of the cystic duct is oriented anteriorly (Fig. 6); this suggests that this duct is elongated and that the gallbladder delivers its secretions close to the stomach in the digestive tract. The kidneys are present on the dorsal margin of the abdominal cavity; they are paired and independent elongated organs anteriorly almost in contact with the occipital region of the neurocranium (Fig. 6).
In angler, the location of the heart is quite unusual; this organ lies in a very anterior position, ventral to the rostral extremity of the skull (Figs. 8 and 9). The atrium is displaced to the left side and is as large as the ventricle (Fig. 8). Cavities full of liquid are rare and not expanded in angler (Figs. 8 and 9); some are present around the intestine coils. The swimbladder is absent (Figs. 8 and 9), the stomach is distinct, well developed and composed of two sequential pouches (Figs. 8 and 9). A long and coiled intestine occupies the main portion of the abdominal cavity. These coils are horizontally organized, with two short pyloric caeca (Fig. 8). In our images the organisation of the digestive tract is similar to that seen in Home's work (Fig. 10) [12]. The liver is composed of two lobes, with the left one much more developed than the right one (Fig. 8). The gallbladder is a well developed rounded structure present on the right side, between the two hepatic lobes. Its opening is oriented anteriorly (Fig. 8), suggesting that the opening of the bile duct in the digestive tract is close to the stomach and that the cystic duct is elongated. The brain occupies only a very small part of the cranial cavity (Fig. 9A); it is surrounded by a tissue appearing in white in T2. The kidneys are present in the dorsal margin of the abdominal cavity; they are paired, independent and rounded organs not extending anteriorly to the neurocranium (Fig. 9B).
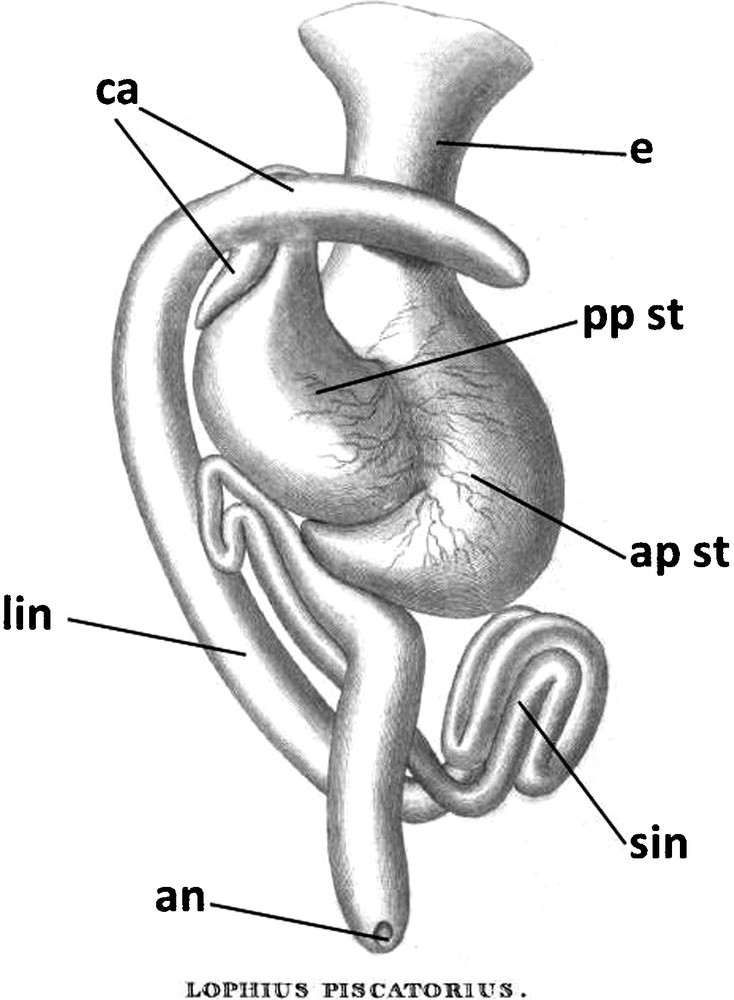
Organization of the digestive tract of angler, drawing present in [12]. an: anus; ap st: anterior pouch of the stomach; ca: pyloric caeca; e: esophagus; lin: large intestine; pp st: posterior pouch of the stomach; sin: small intestine.
Modified from plate XCIV [12].
4 Discussion
Previous works dedicated to the anatomy of soft organs of these species are dated and deal with only selected regions of the viscera. Non-invasive studies using imaging techniques are important in the study of the internal anatomy of rare specimens. They enable the examination of the entire visceral anatomy without destroying any of the other organs [42–50]. Moreover, with rarely caught species (such as ocean sunfish), or expensive species (such as angler, around 25 euros per kg at French market auctions in June 2012), the non-invasiveness of these techniques preserves the specimens for future traditional dissections [46–48].
The gross anatomy of these species has been broadly known since the beginning of the 20th century [11–17], with some insights to the detailed anatomy of some regions [16,51]. The absence of a swimbladder in ocean sunfish and angler has already been mentioned [1,2,11,16]. Cuvier briefly described the digestive tract of the angler and the liver of the ocean sunfish [11,13]. Home illustrated the anatomy of angler [12] (Fig. 10). Jones illustrated the cardiac organisation of the angler [14] while quite complete descriptions of the visceral organisation of ocean sunfish have also been published [15,16]. Curiously, there is no indication of the sizes of the animals or the organs in any of these works (Fig. 10). Heart and digestive tracts were apparently taken out before description (Fig. 10).
Consequently, the occurrence of cavities full of liquid in ocean sunfish has been rarely mentioned before. Only Lacépède briefly mentioned them [52], and the presence of such cavities is quite unusual among vertebrates; MRI examinations of the anatomy of some other teleostean fishes did not show similar structures (see the numerous images in the DFL). Even if the occurrence of similar chambers has to be confirmed in other species, we may hypothesize that these cavities are important to ocean sunfishes to compensate the effects of pressure while diving frequently from the surface to depths of several hundred meters [2].
The present examinations show that both species lack a swimbladder, have an asymmetry in the hepatic lobes, possess a bile duct that terminates close to the stomach, have independent kidneys and a very reduced brain (Table 1). The thyroid gland is not visible on the MRI images of angler, while this gland appears to be visible on the MRI image of the ocean sunfish. The structure of this gland, a full of liquid pouch next to the ventral aorta, is similar to the description of the thyroid for the angler [51]. Such an organization of this endocrine gland is quite peculiar among teleosteans, where this gland is usually diffuse [53,54], and is present as well in Canthidermis rotundatus, Balistidae [55] and Diodon holacanthus, Diodontidae [56]. A swimbladder is present in more basal tetraodontiform species, as D. holacanthus, where this organ is involved in the well known process of inflation [57]. The absence of a swimbladder in angler and ocean sunfish may be explained by the ecology of the animals [1–5,7]. Angler is a sedentary bottom-dwelling species living at 30-500 m [5,7] and the absence of a swimbladder is viewed as an adaptation to life on the bottom of the sea in this species [2]. Ocean sunfishes are active swimmers able to dive 40 times a day from the surface to a depth of 600 m [2]. A swimbladder might hamper, or even preclude, this diving behaviour; this organ is also absent in some continuously swimming teleosts like Atlantic mackerel (Scomber scombrus, Scombridae) or little tunny (Euthynnus alletteratus, Scombridae), two species capable of fast acceleration [2]. A very asymmetric liver, a bile duct opening close to the stomach and a reduced brain (remarkable among teleostean species) have not been described in other acanthomorph species and is not mentioned in basal tetraodontiform species. Independent kidneys have been described in the angler [58]. The presence of an asymmetric liver, a long cystic duct opening close to the stomach, independent kidneys and a compact thyroid gland might be considered as probable synapomorphies of these two species and may confirm the close relationships of Tetraodontiformes and Lophiiformes not yet corroborated by other anatomical data. These characters, however, have to be observed in more species before integrating them in a data matrix.
Similitude and differences in the visceral anatomy between angler (L. piscatorius) and ocean sunfish (M. mola).
| Lophius piscatorius | Mola mola | |
| Brain | Reduced | Reduced |
| Spinal cord | Not extending far in the spinal canal [18] | Not extending far in the spinal canal [17] |
| Heart | Present very anteriorly position, ventral to the rostral extremity of the skull. Not embedded in an expanded and full of liquid pericardial cavity | Not situated anteriorly, voluminous and in an expanded and full of liquid pericardial cavity |
| Thyroid gland | Compact in a blood lacuna [48] | Compact in a blood lacuna |
| Kidneys | Paired, independent and rounded organs [55] | Paired and independent elongated organs |
| Swimbladder | Absent | Absent |
| Structure of the liver | Asymmetric with a more developed left lobe | Asymmetric with a more developed left lobe |
| Gallbladder | Developed with a long cystic duct opening anteriorly, close to the stomach | Developed with a long cystic duct opening anteriorly close to the stomach |
| Digestive tract | Coiled with two pyloric caeca | Deeply coiled with no pyloric caecum |
Non-invasive imaging techniques, as used here, appear to be important in studying and deciphering the details of the anatomy of species. In the Digital Fish Library (DFL), images of the molid genera Mola, Masturus, and Ranzania and the lophiiform genus Lophoides (Lophiidae) are available. These images have been examined. Unfortunately, differences exist in the resolution between these images (Fig. 2). The resolution of MRI images depends on several factors such as the size of the specimen, the field of view, magnet strength, gradient strength, and the type of protocol [45,48]. But all MRI parameters are not provided in the DFL, such as weighting parameters (T1 versus T2), although recent work [46–49] has shown the importance of mentioning these parameters in recognizing internal structures, and especially in identifying presence of liquid inside the organs. MRI images present in [50] are of high quality, but even if they represent a definitive input for comparative anatomy studies, all the images present in the DFL do not present such a resolution (Fig. 2) and therefore cannot be used here for anatomical comparisons between angler and ocean sunfish.
Despite the fact that ocean sunfish and angler have been known since ancient times and have been the subject of many publications, the internal anatomy of these animals has been rarely described and had never been compared. Even if they greatly differ in their external anatomy, the present study shows that angler and ocean sunfish present some similarities in their internal anatomy (Table 1). Therefore, before explaining these similarities by common ancestry, the internal anatomy has to be examined in other related species, such as frogfishes (Antennariidae), boarfishes (Caproidae), triggerfishes (Balistidae), puffers (Tetraodontidae) or boxfishes (Ostraciidae and Aracanidae). Unfortunately, MRI examinations with a 1 to 2 Tesla magnetic fields do not allow examining such specimens. Most puffers, frogfishes and boxfishes are quite small; previous works [46] showed that anatomical studies using 1 to 2 Tesla MRI cannot be successfully conducted on specimens smaller than 15 cm in total length. Nevertheless, non-invasive imaging techniques appear here as efficient tools to investigate the internal anatomy of rare, valuable and large animals, before conducting dissections on such animals as ocean sunfish and angler. The internal anatomy of smaller animal, as Drosophila has been successfully investigated in vivo by MRI at 18.8 Tesla [59]. We hope that future studies using MRI coupled with CT imaging on smaller specimens, as well as classical dissections, will continue to employ this approach.
Disclosure of interest
The authors declare that they have no conflicts of interest concerning this article.
Acknowledgments
We express our deepest gratitude to Dr. A. Grenouilloux (Rennes, France), Pr. J.-C. Desfontis (Oniris, Nantes, France) and C. Lenglin (Aquarium, Granville, France) for support, to F. Chapleau (Université d’Ottawa, Ontario, Canada) for bibliographic help, to J.F. Dejouannet (UMR 7138 CNRS and pole dessin UMS 2700 CNRS, MNHN, Paris, France) for illustrations, to M. Comte, P. Bugnon, C. Costes, C. Picard, F. Lebatard and K. Harfoush (Oniris, Nantes, France), J.-M. Léger (Leclerc, Nantes, France) and C. Cauchie (Douai, France) for technical support and to I. Nicholson (Oniris, Nantes, France) for improvements of the manuscript. The authors thank Prof. L. Frank, Dr. R. Berquist and staff at the Digital Fish Library (www.digitalfishlibrary.org) for providing MRI images for M. mola and the Fondation TOTAL for support. We are grateful to Prof. J.T. Eastman (Department of Biomedical Sciences, Ohio University, USA) who reviewed carefully our text. We thank the two anonymous reviewers for their acute and supportive comments on first versions of the manuscript. This work was supported by the Fondation TOTAL (project “Classification des poissons marins: les téléostéens acanthomorphes”).