

1 Introduction
Mycorrhizae are symbiotic associations between plant roots with some fungi soil. Mycorrhizae directly influence mineral nutrition of the plant, water absorption, and resistance to biotic and abiotic stresses [1–4]. The plant generally provides carbohydrates to the fungus. Exchanges between the two hosts may vary according to the type of present mycorrhizal symbiosis [5]. Arbuscular endomycorrhizal fungi belong to the phylum of Glomeromycota [6]. They can develop arbuscules and vesicles [7]. These fungi have very little specificity to the host plant. In fact, 130 species of these fungi ensure mycorhization of more than 300,000 plant species [5].
It has been shown that various ligneous tree are able to develop a symbiosis with endomycorrhizal fungi species, such as marula (Sclerocarya birrea) and argan (Argania spinosa). Studies showed that the inoculation of different varieties of marula by 250 spores Gigaspora margarita has allowed one to obtain mycorrhizal young plants at a rate varying by around 40% [8]. Mycorrhization largely improved the resistance of these fruit tree species to water stress, salt well, as well as to flooding conditions. Other research showed that the argan plants colonized with a rate of 70% after their inoculation with different strains of the Glomus genus [9]. This mycorhization permitted a gain in dry biomass of aerial and root parts, respectively equal to 120% and 70%, and improved nutrition elements, such as phosphorus, potassium, calcium, manganese, and copper. Inoculation of the same tree species presented an index of relative mycorrhizal dependency (IRMD) equal to 48% [10]. Jacaranda mimosifolia is also a ligneous tree. It is therefore possible that it withdraws benefits if it is mycorrhized at its nursery production, especially as these mycorrhizal inoculants are commercially available or can be trapped and multiplied under controlled conditions and with mycotrophic plants [11,12]. This requires a better understanding of various characteristics between endomycorrhizal fungi and ornamental ligneous tree, as well as possible interactions between inoculum and fertilizers doses. Indeed, these parameters can affect the level of mycorhization and, consequently, the development of the plant.
The aim of this study is to assess the mycorrhizal status of Jacaranda mimosifolia D.Don growing in nurseries, after various arbuscular endomycorrhizal inoculations in the presence of different levels of fertilization.
2 Materials and methods
2.1 Vegetal material and treatments applied
One-year young 40 cm high Jacaranda plants that were obtained using seedlings were inoculated with two inoculums of different origins and compositions, according to different modes of application, and fertilized with increasing doses. These inoculants and fertilizer intakes were made simultaneously when repotting young plants of Jacaranda in 10/12 cm size pots, containing a substrate composed of peat and sand, with proportions respectively equal to 2/3 and1/3. The experiment was conducted in a greenhouse at the “Institut national agronomique de Tunisie” under homogeneous conditions; the temperature was maintained constant at 25 °C. After six months of cultivation, mycorhization rates of roots that had undergone different Jacaranda treatments were evaluated.
2.2 Fertilizer applied
For each treatment of inoculum, four fertilizer levels were tested. Three repetitions were performed for each combination of inoculum and fertilizer as a treatment. The fertilizer used is the Osmocote EXACT standard-Scotts, a slow release NPK fertilizer (5–6 months), containing magnesium and trace elements 15 + 9 + 12 (+ 2.5). The detailed composition of the fertilizer is presented in Table 1. The tested doses of fertilizer and their corresponding proportions of nitrogen, phosphorus, and potassium are displayed in Table 1.
Composition of fertilizer Osmocote EXACT.
| Element | Concentration (%) |
| Nitrate nitrogen | 7 |
| Ammoniacal nitrogen | 8 |
| Phosphoric anhydride (P2O5) | 9 soluble in neutral ammonium citrate and in water; 6.8 soluble in water |
| Potassium oxide (K2O) | 12 soluble in water |
| Magnesium oxide (MgO) | 2.5 and 1.3 soluble in water |
| Bore (B) | 0.02 soluble in water |
| Copper (Cu) | 0.068 and 0.051 soluble in water |
| Total iron (Fe) | 0.45 |
| Iron soluble in water | 0.31 |
| Iron chelated by EDTA | 0.22 |
| Manganese (Mn) | 0.06 and 0.03% soluble in water |
| Molybdenum (Mo) | 0.025 soluble in water |
| Zinc (Zn) | 0.02 and 0.013 soluble in water |
2.3 Used inoculums
The inoculums used are made of arbuscular endomycorrhizal fungi and differ in their origins, nature and methods of application.
The first tested inoculum is in the form of a substrate containing spores and roots colonized by five strains from the genus of Glomus : Glomus constricticum, Glomus geosporum, Glomus fuegianum, Glomus irregulare and Glomus sp. These indigenous arbuscular endomycorrhizal fungi were trapped from soils of three prospected sites of Nahli National Park (Tunisia), on a substrate composed of one volume of perlite, four volumes of clay granule (Terra green) and one volume of vermiculite [13]. Amplification of the inoculum was then conducted by greenhouse crops of leek (Allium porrum L.) and vetch (Vicia sativa L.) during four months on a poor nutrient substrate made with sand and perlite with variable proportions of 2/3 and 1/3, respectively. Application of this inoculum was carried out in terms of percentage of the pot's volume. Two doses have thus been prepared:
- • dose 1: 10% inoculum + 90% substrate (2/3 peat + 1/3 sand);
- • dose 2: 20% inoculum + 80% substrate (2/3 peat + 1/3 sand).
The second endomycorrhizal inoculum tested is a commercial product. It belongs to the brand MYKE PRO, which is produced by Premier Tech Itée, Rivière-du-Loup, QC, Canada. This product contains G. irregulare as an active ingredient, with a proportion of 1600 propagules/g, a density of 500 g/L and a granulometry of 0.5 mm. The inoculum bag was stored at 4 °C until use. Doses are expressed in grams, namely 0.06 g and 0.1 g, which were applied for each plant within the pot (Table 2).
Proportions in nitrogen (N03), phosphorus (P2O5) and potassium (K2O) sources corresponding to the applied dose of fertilizer.
| Dose of fertilizer Osmocote Exact (g/plant) | 0 | 2 | 4 | 6 |
| Quantity brought of nitrogen source (NO3:mg/plant) | 0 | 300 | 600 | 900 |
| Quantity brought of phosphorus source (P2O5:mg/plant) | 0 | 180 | 360 | 540 |
| Quantity brought of potassium source (K2O:mg/plant) | 0 | 240 | 480 | 720 |
2.4 Evaluation of endomycorrhizal colonization
Root fragments are randomly selected at various levels of the underground parts of the different plants for each treatment. Thinning using KOH (10%) and oxidation of the organic matter, which is present on roots, was carried out firstly to eliminate intracellular components and leave only the root and fungal structures. These roots are colored by fushine acid (0.05% in lactoglycerol). Colored roots are preserved in lactoglycerol (25% lactic acid, 25% glycerol, 50% water) to dilute the unfixed colorant and prevent drying. The fushine acid is generally used to color selectively dead tissues or cells. It is, indeed, a colorant capable to be fixed on the chitin of all the fungal structures. A root is considered as endomycorrhized when it presents an endomycorrhizal structure (mycelium, vesicle, and arbuscule) [14]. The evaluation of the rate of roots mycorrhization by endomycorrhizal fungi is achieved through the method of McGonigle and Fitter [15]. For the quantification of endomycorrhizae, microscopic examination (40 to 200 × ) of 80 1-cm long root fragments was performed for each plant. The root fragments were randomly selected and mounted in parallel by groups of 10 between the slide and cover. Three readings per fragment were conducted to determine the type of the fungus structure.
Each experimental unit included three pots in which one plant was assessed. Since all the assessments were carried out within a green house, it was assumed that all experimental units were homogeneous, and a completely randomized experimental design was used according to the following model:
Experiments with both indigenous composite inoculum and commercial inoculum G. irregulare were performed simultaneously and under the same environmental conditions.
3 Results
3.1 Mycorrhizal colonization of Jacaranda roots in the presence of five indigenous strains of Glomus genus present within the substrate
Microscopic observations of the colored roots of Jacaranda indicate that all plants having received a proportion of indigenous composite inoculum were mycorrhized, for all tested fertilization levels. Fungal structures found in the root tissues are essentially hyphae with fungal filaments bobbins inside certain cells (Fig. 1).
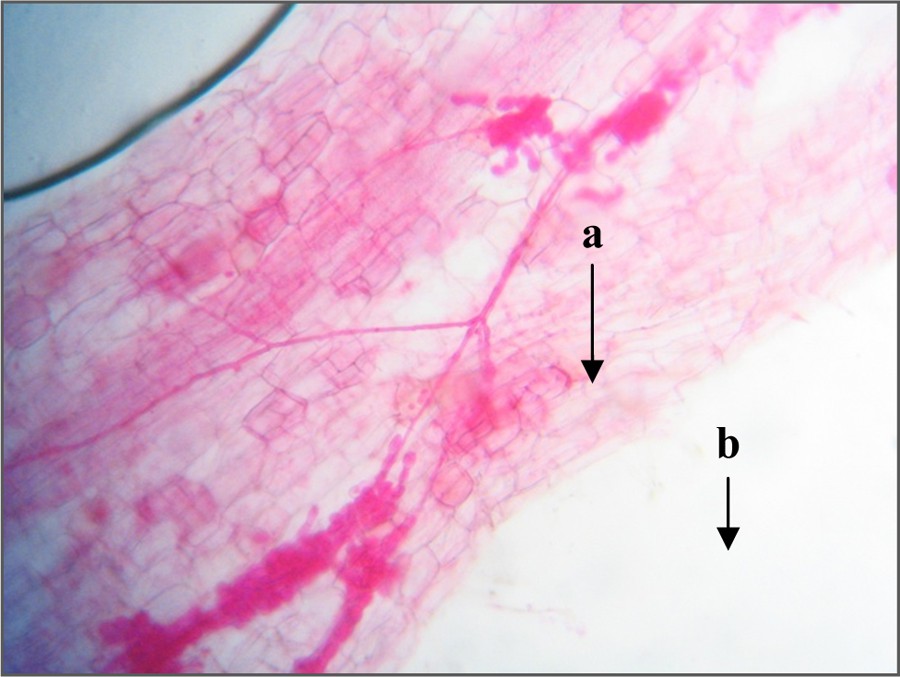
Arbuscular endomycorrhizal structures observed on the roots of young plants of Jacaranda mimosifolia D.Don, after six months of composite indigenous inoculum intake (400× magnified images); a: hyphae; b: coils fungal filaments within cells of root fragment. Color online.
The study of the effect of the interaction between the fertilizer dose and the inoculum proportion contained in the pot on the Jacaranda root colonization rate (Table 3) shows that it is highly significant (P < 0.01). However, according to Fig. 2, which shows the root colonization rate of Jacaranda plants according to the brought doses of fertilizer and the proportions of composite indigenous inoculums, some heterogeneity is noted concerning the root colonization rate compared to the proportion of inoculum contained in pots. Thus, the recorded rates of colonization evolve independently of the brought proportions of this inoculum. In fact, regarding plants receiving doses of 0 g and 4 g of fertilizer, the most important endomycorrhizal colonization was observed on the roots of Jacaranda cultivated on substrates containing only 10% of the indigenous inoculum. On the one hand, the colonization rate of unfertilized plants is 57.1% for plants grown on a substrate containing 10% of the indigenous inoculum, while it did not exceed 37.85% for those grown on a substrate containing 20% of the inoculum. On the other hand, for fertilization with 4 g of Osmocote/plant, the colonization rates are 31.29% and 8.64%, respectively, for plants grown on substrates containing 10% and 20% of the indigenous inoculums. However, in case of a fertilization dose of 2 g/plant, the highest colonization rate was recorded with plants receiving the highest dose of inoculum. This rate was equal to 32.87% for an inoculum of 20%, while it was only 6.66% for a 10% inoculum. Finally, for the highest level of fertilization, namely 6 g of fertilizer/plant, the recorded colonization rates with both tested doses of inoculum are low (less than 3%). Therefore, the possibility of the existence of an interaction effect between the dose of fertilizer and the proportion of indigenous composite inoculum on the level of mycorhization in Jacaranda was excluded.
Effect of variation in the brought proportion of composite indigenous inoculum and fertilizer dose on root colonization rates of Jacaranda mimosifolia D.Don.
| Inoculum proportion contained in the total volume of the pot (percentage of the total volume of the pot) | Fertilizer dose (g) | Colonization rate (%) |
| 10 | 0 | 57.10a |
| 20 | 0 | 37.85b |
| 20 | 2 | 32.88b,c |
| 10 | 4 | 31.3c |
| 20 | 4 | 8.64d |
| 10 | 2 | 6.66d,e |
| 10 | 6 | 2.59e |
| 20 | 6 | 2.22e |
| F trait | ** | |
| Variation coefficient | 14.32 | |
| R 2 | 0.99 |
** Highly significant difference between treatments (P = 0.01).
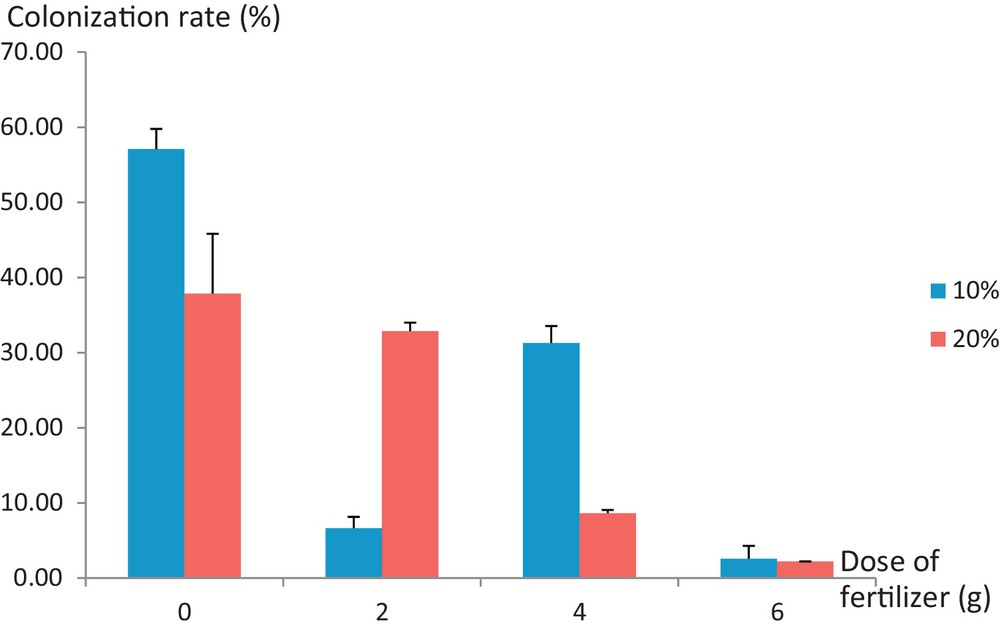
Mycorrhizal colonization (%) based on the doses brought into fertilizer and inoculum indigenous composite proportions contained in pots; 10%, 20%: inoculum proportions expressed in terms of volume percentage of the pot. Color online.
The root colonization rate of Jacaranda plants varies significantly (Table 4) according to the level of fertilization (P < 0.01). Root dry biomasses of treatments vary in the same way as the rates of root colonization, depending on the levels of fertilization (P < 0.01).
Effect of the fertilization level on root colonization rates of Jacaranda mimosifolia D.Don inoculated with two inoculums with different origins, natures, and intake modes.
| Dose of fertilizer (g) | Composite indigenous inoculum | Commercial inoculum | ||
| Colonization rate (%) | Root dry mass (g) | Colonization rate (%) | Root dry mass (g) | |
| No fertilization | 47.48a | 4.35a | 32.35a | 4.18a |
| 2 | 19.77b | 2.72c,b | 15.82b | 2.14b |
| 4 | 19.97b | 2.99b | 17.12b | 1.80c |
| 6 | 2.41c | 2.66c | 3.95c | 2.32b |
| F trait | ** | ** | ** | ** |
| Variation coefficient | 14.33 | 8.19 | 44.73 | 9.28 |
| R 2 | 0.99 | 0.96 | 0.78 | 0.97 |
** Highly significant difference between treatments (P = 0.01).
Indeed, the highest and lowest root colonization rates and root dry biomasses are respectively observed in unfertilized and fertilized 6 g/plant treatments. With 2 g and 4 g of fertilizer per plant, the root colonization rates were not significantly different between treatments and root dry biomasses were very similar (Table 4). Generally, the rate of root colonization by indigenous endomycorrhizal complex decreases with increasing the fertilizer dose. Dry root biomasses of treatments vary depending on their levels of mycorhization.
3.2 Mycorrhizal root colonization of Jacaranda in the presence of a commercial inoculum G. irregulare in the substrate
Microscopic observations of Jacaranda colored roots treated with commercial inoculum show that at all levels of fertilization, all the plants are mycorrhized. Arbuscular endomycorrhizal structures observed on the roots are essentially in the form of hyphae with a limited presence of exchange (arbuscules) and reserve (vesicles) structures. Fig. 3 shows a hypha penetrating into a root cell and forming a ramification that constitutes the arbuscule.
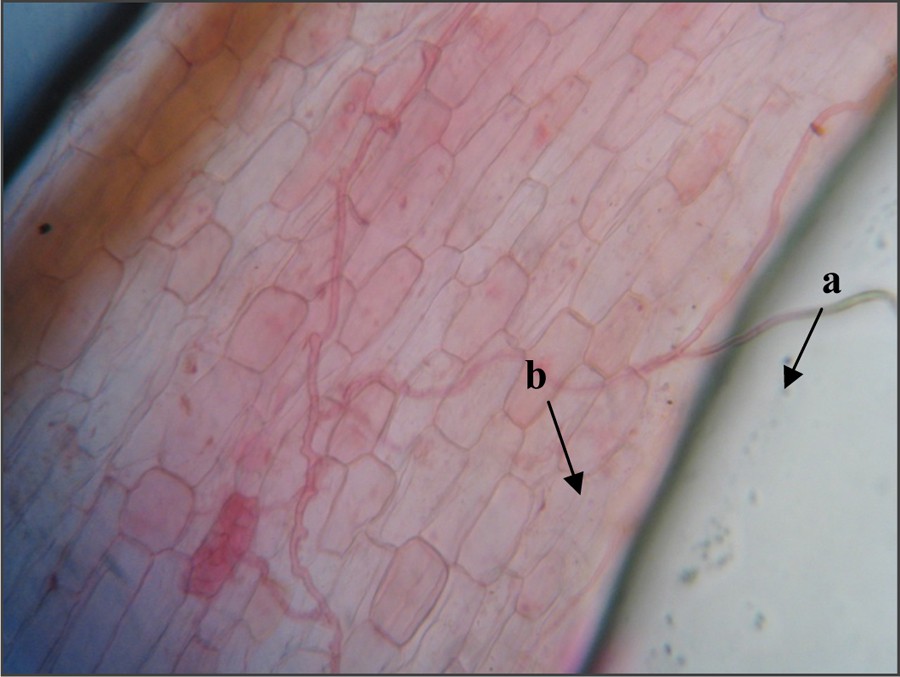
Observed structures on a root fragment of Jacaranda mimosifolia D.Don, six months after inoculation with a commercial inoculum containing Glomus irregulare (400× magnified image) structures; a: hypha entering a root cell; b: arbuscule. Color online.
Interaction effects between the doses of fertilizer and those of commercial inoculum on Jacaranda root colonization rate have proved insignificant in this study. Parameters “inoculum dose” and “level of fertilization” were then studied as having simple effects on the rate of mycorhization.
The rates of root colonization are evolving positively based on increasing brought doses of inoculum (Fig. 4). For all levels of fertilization, the most mycorrhized Jacaranda plants are those that received the highest dose of inoculum, i.e. 0.1 g/plant. The effect of the inoculum dose on the rate of root colonization of young Jacaranda plants (Table 5) was significant (P < 0.05). Providing a larger dose in commercial inoculum, namely 0.1 g/plant, an increase of 42.1% in the root colonization rate of Jacaranda was observed compared to the situation with a 0.06 g/plant dose.
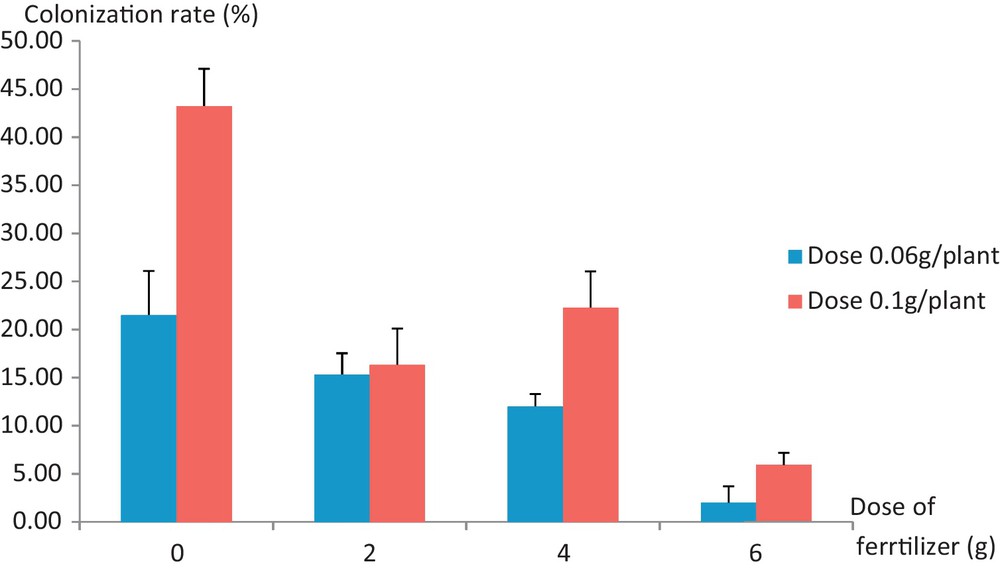
Colonization rates (%) according to the brought fertilizer and commercial inoculum doses. 0.06 g/plant, 0.1 g/plant: industrial inoculum doses. Color online.
Effect of the brought dose of commercial inoculum on root colonization rates of Jacaranda mimosifolia D.Don.
| Inoculum dose (g) | Colonization rate (%) |
| 0.06 | 12.69b |
| 0.1 | 21.92a |
| F trait | * |
| Variation coefficient | 44.73 |
| R 2 | 0.78 |
* Significant difference between treatments (P = 0.05).
The rate of root colonization varies significantly based on the level of fertilization (P < 0.01). Thus, it appears that increasing the brought fertilizer dose decreases the level of mycorrhizal Jacaranda plants treated with commercial inoculum (Table 4). Indeed, the highest and lowest rates of root colonization are respectively observed in unfertilized samples and in those that received 6 g of fertilizer/plant. The rates root colonization treatments with doses of 2 g and 4 g/plant of fertilizer are not significantly different. Root dry biomasses do not evolve exactly as the level of mycorhization. However, non-fertilized but better mycorrhized plants have significantly higher root dry biomasses (P < 0.01) than the rest of the sample (Table 4).
4 Discussion
4.1 Quality of root colonization of young Jacaranda plants treated by different inoculums
According to the obtained results, the plants treated with composite indigenous inoculum reached a more advanced stage of mycorrhization than those treated with the commercial inoculum. Indeed, microscopic observations of the latter showed only small colony dispersed on the mycorrhized roots instead of fungal structures extending over the entire lengths. These roots display rare arbuscules. In addition, the vesicles are almost non-existing, indicating a recent colonization. On the contrary, the presence of fungal filaments coils on the roots of Jacaranda plants treated with composite indigenous inoculum reflects a more advanced stage of mycorrhization. In fact, these coils represent structures that appear in Paris-type colonization. The colonization of the root by hyphae of arbuscular mycorrhizal (AM) fungi can adopt two morphological types: the Paris and the Arum ones [16]. Colonization of Arum type has an extensive intercellular evolution of fungal hyphae in the root cortex, with the development of intracellular arbuscules, while colonization of Paris-type is defined by the absence of intercellular phase and the presence of numerous bobbins of intracellular hyphae. Arbuscules are structures that intercalate between the coils [16]. In case of such colonization, the arbuscules are relatively few, small, or even non-existing [16]. Nevertheless, this shows that colonization of Jacaranda plants treated with composite indigenous inoculum was well established.
4.2 Effect of the variation of the inoculum brought dose on the mycorrhizal level of young Jacaranda plants
The existence of an interaction effect between inoculum dose and the level of fertilization on the rate of mycorhization in Jacaranda was excluded. These two variation parameters were studied as simple effects.
The obtained results showed that the level of Jacaranda's mycorhization is closely related to the variation of the brought dose of commercial inoculum. The highest rates of colonization were observed in plants treated with 0.1 g. These results corroborate with the other studies of the tested isolates of Glomus sp. on pineapple vitroplants. Their results showed that an increase in inoculum doses from 1% to 3% was often linked to a rise in the endomycorrhizal infection [11]. Contrary to Jacaranda plants treated with composite indigenous inoculum, the levels of mycorrhization have evolved independently of the variation in the proportion brought. These different results can be explained by the nature of the used inoculum. In fact, commercial inoculum is a homogeneous product, concentrated in propagules and adapted according to the application, while the composite indigenous inoculum is a product of trapping and multiplication with different mycotrophic plants. It is made of propagules of arbuscular endomycorrhizal fungi and mycorrhizal root fragments mixed in an inert substrate. The latter may present some heterogeneity in the concentrations of propagules and mycorrhized root fragments. The proportions applied to Jacaranda plants may not contain the same quantities of inoculants structures, which could explain the random changes in the levels of mycorhization after varying the proportions brought in this inoculum.
4.3 Effect of the level of fertilization variation on the mycorhization rate of young Jacaranda plants
The effect of varying the dose of fertilizer on the mycorhization level of young jacaranda plants has proved highly significant (P = 0.01) in both types of inoculum. In fact, increasing the brought dose of fertilizer reduced the infectivity of arbuscular endomycorrhizal fungi contained in the inoculum, thus, inducing a decrease in the root colonization rate of Jacaranda plants. These results corroborate those obtained with the studies on the interactions between extractible phosphorus soil level and colonization by arbuscular mycorrhizal fungi of white clover (Trifolium repens L.) [17]. The level of root colonization was negatively correlated with extractible phosphorus soil concentration. [17]. Studies on the effect of fertilization at different levels of phosphorus and nitrogen on mycorhization of Allium shoenoprasum with G. caledonium showed that the simultaneous application of high doses of phosphorus and nitrogen led to a decrease in the rate of root colonization. [18]. This effect is more apparent with ammonia's nitrogen than with its nitrate form [18].
Therefore, it is noticed that a too high level of mineral elements in soil inhibits the formation of mycorrhizae. In fact, increasing nitrogen and phosphorus concentration in the soil solution promotes the synthesis of proteins and of phosphorylated compounds (nucleic acids, DNA and RNA, inositol, phosphates, etc.), which causes a decrease in the soluble sugar content in the root. Since the soluble sugar content of the root is a key parameter for the nutrition of mycorrhizal fungi associated with the plant, it also determines the rate of formation of mycorrhizae [19]. According to these results, mycorhization has a positive effect on root dry biomass of Jacaranda, as in the case of unfertilized plants. Intake of different doses of fertilizers did not allow them to reach such a level of root system development. These results corroborate other studies on the role of arbuscular mycorhization in the improvement of the morphology and the development of host plant root system, e.g., those reporting the inoculation of “Annona cherimola Mill” with propagules of arbuscular mycorrhizae, which allowed a significant increase in the branching intensity of the first-order laterals [20].
5 Conclusion
At all the levels of fertilization, plants of Jacaranda having received a dose of inoculum were mycorrhized. Six months after inoculation, the plants treated with the composite indigenous inoculum reached a more advanced stage of mycorhization than those treated with the commercial inoculum.
For Jacaranda plants inoculated with the commercial product, the variation in the level of mycorhization has proved to be closely related to the inoculum brought dose. Then, an improvement in the rate of root colonization by arbuscular endomycorrhizal fungi with an increase in the inoculum dose is observed. For plants treated with different proportions of composite indigenous inoculum, the level of mycorhization varies randomly. This can be explained by the existence of some heterogeneity of the concentrations of the propagules contained in the brought inoculum proportions.
For the two types of inoculum, a too high level of fertilization inhibited the infectivity of arbuscular endomycorrhizal fungi, then, inducing few mycorrhized Jacaranda plants. The best root colonization rates were observed in the unfertilized plants, which have the most important root dry biomasses.
Disclosure of interest
The authors declare that they have no conflicts of interest concerning this article.