

1 Introduction
Diplometopon zarudnyi (D. zarudnyi), commonly kwon as Zarudny's worm lizard, is a species of amphisbaenian reptile in the Trogonophidae family. It is monotypic within the genus Diplometopon [1]. This species is generally found in some pockets of the Arabian Peninsula such as west Iran, south Iraq, Kuwait, Oman, United Arab Emirates, and north Saudi Arabia [2,3]. D. zarudnyi is an oscillating digger, commonly found burrowing in low sand dunes in open terrain and in sub-surface soils of date palm farms. This amphisbaenian is a nocturnal forager, sometimes occupying ant and termite tunnels.
In recent years, several studies were published on spermiogenesis of reptiles such as Agama adramitana [4,5], freshwater turtle Maurymes caspica [6,7], Acanthodactylus boskinus [8]; House Gecko (Ptyodactylus hasselquisti) [9], and Jamaican Gray Anole (Anolis lineatopus) [10]; however, till date, there is no data on spermiogenesis in D. zarudnyi (a worm lizard). Hence, there is a need to examine the process of head differentiation, which enables us to compare a reptile with that of other vertebrates. Therefore, an attempt has been made to explore the ultrastructural differentiation of the head region in D. zarudnyi. The outcome of this study provides the relationship between the phylogenetic aspects of other reptiles.
2 Materials and methods
2.1 Animal collection
Five adult male Zarudny's worm lizards (D. zarudnyi) were collected from April to May 2011 in the Riyadh province, specifically from the Thummama and Dilam regions.
2.2 Histological study
Soon after sacrifice, the testis was fixed in 10% buffer formalin and was subsequently preserved in 70% alcohol. Histological sections were made at intervals of 4 μm and stained in haematoxylin and eosin.
2.3 Ultrastructural study
Immediately after removal of the testis from the dissected lizards, tissues were sliced into small size samples (1 mm3) and fixed in 3% buffered glutaraldehyde for 4 h at 4 °C. Tissue specimens were then post fixed in 1% osmium tetraoxide (OsO4) for 90 min. Dehydration of the fixed tissue was performed using ascending grades of ethanol and then tissue were transferred into the resin via propylene oxide. After impregnation with pure resin tissue, specimens were embedded in the same resin mixture [11]. Ultra-thin sections of silver shades (60–70 nm) were cut using an ultra-microtome (Leica, UCT) equipped with a diamond knife; sections were then placed on copper grids and stained with uranyl acetate (20 min) and lead citrate (5 min). Stained sections were observed with a TEM (JEOL JEM-1011) operating at 80 kV.
3 Results
The initiation of spermiogenesis is marked by the accumulation of round spermatids within the seminiferous epithelium of D. zarudnyi upon the completion of meiosis. In Fig. 1, an overall morphology of the sperm was observed using a light-microscope. During the ultrastructural study, the proacrosomal vesicle develops from the Golgi complex that was adjacent to the nucleus (Fig. 2). It is then pressed against the nuclear wall in which it forms a wide shallow depression. As the proacrosomal vesicle advances, a dense granule appears in its interior (Fig. 3). This granule is initially attached to the vesicle membrane, in contact with the nucleus. Between the nucleus and the vesicle, an electron-dense fibrous layer is observed (Fig. 4). Then, the subacrosomal granule is formed in the middle of the fibrous layer under the acrosomal granule (Fig. 4); it contacts both the acrosomal granule and the outer nuclear membrane. The early spermatid has a round nucleus and the spermatid nucleus moves eccentrically at this stage (Fig. 5).
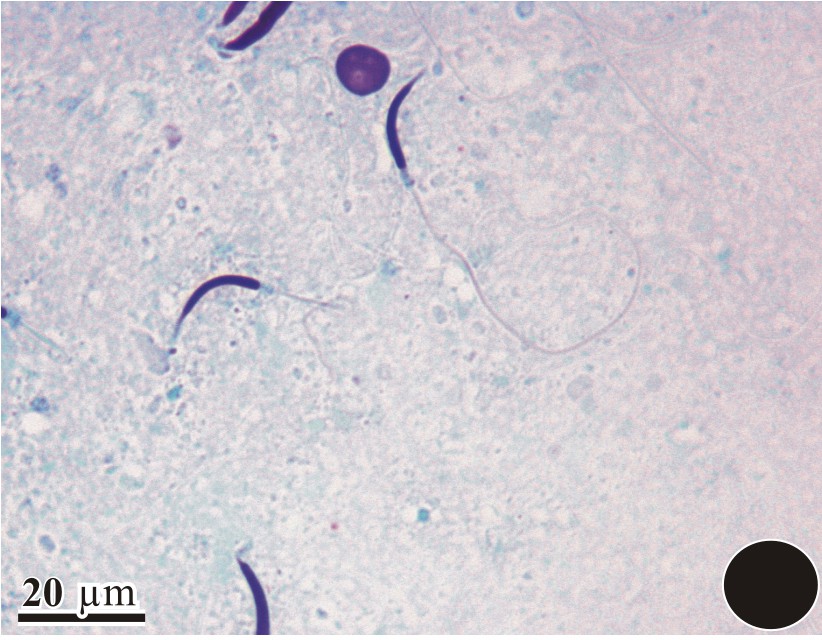
Light-microscope view of a complete sperm showing the curved head and the elongated tail region.
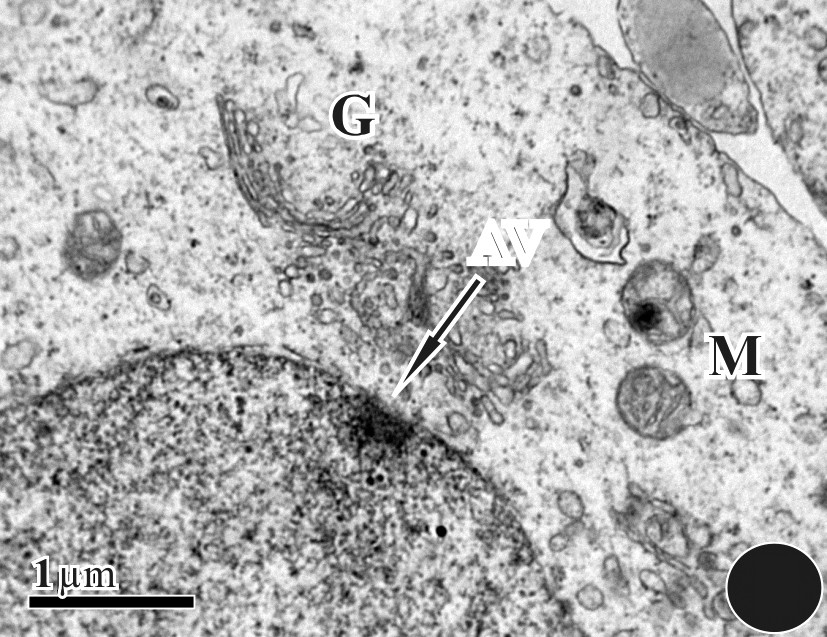
Round spermatid undergoing vesicle (arrow) formation in a region, surrounded by Golgi bodies (G) and mitochondria (M).

Invagination and thickening of the nuclear membrane; formation of the acrosomal vesicle in its early stages.
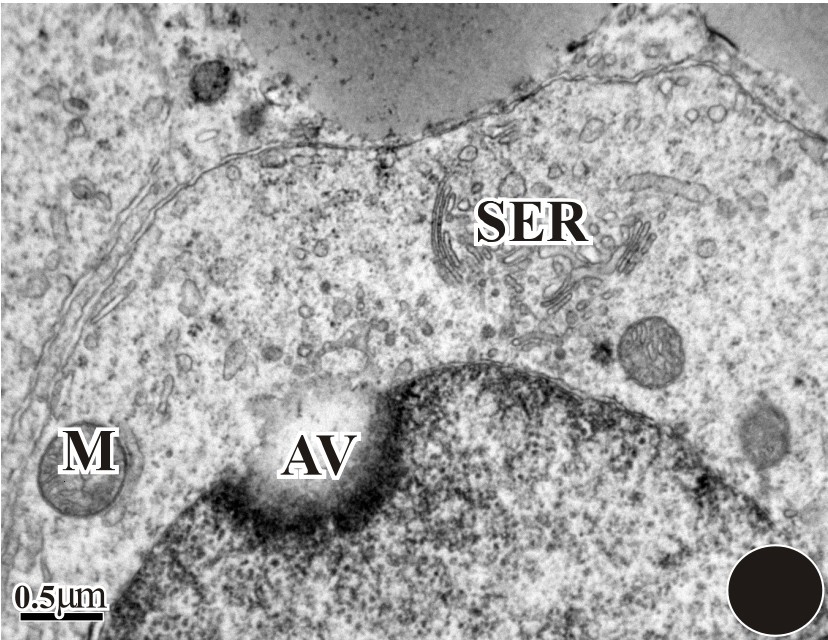
The nuclear membrane becomes thick (dark) with deep invagination and development of the acrosomal vesicle (AV). In the cytoplasm region, mitochondria (M), Golgi bodies (G), and SER were seen.

A round nucleus with a distinct acrosomal vesicle (AV) was seen. The boundaries of the round spermatid were well demarcated.
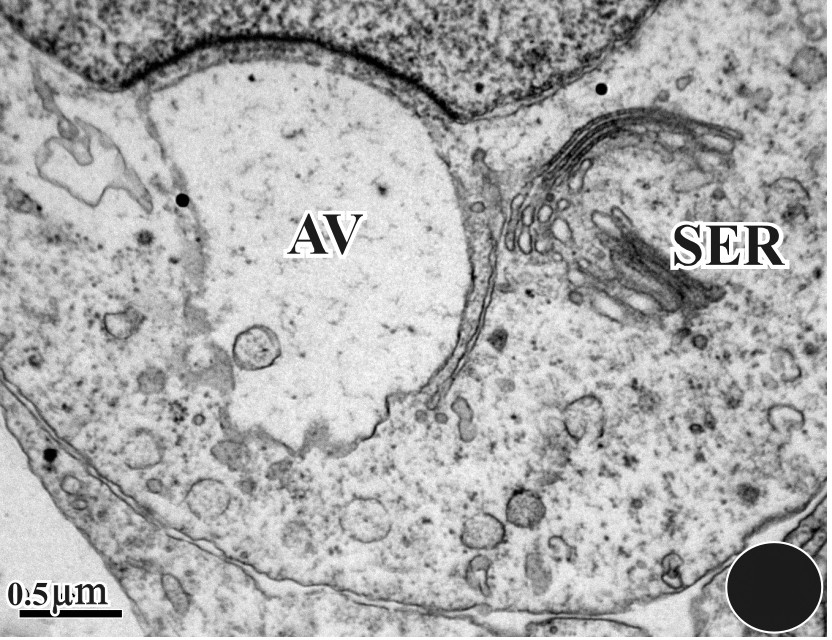
The anterior region of the nucleus forms a protrusion that pushes against the acrosomal vesicle, which becomes progressively concave (Figs. 5–8). The dense acrosomal granule spreads within the vesicle as within the fibrous layer (Figs. 9 and 10) and is transformed into the subacrosomal cone shown in Figs. 11 and 12. The fine granular nuclear substance of the early spermatid is gradually replaced with numbers of coarse granules that appear to form by aggregation of smaller dispersed elements of the chromatin (Fig. 13). The nucleus becomes very compact and the manchette that had appeared during the onset of chromatin condensation retains an intimate association with the nucleus during its transformation (Fig. 13). The acrosomal complex comprising the acrosomal cap and the subacrosomal cone begins to build its final structure, which covers the initial narrowed portion of the nucleus (Figs. 13 and 14). The manchette microtubules are seen to have completely changed their orientation with respect to the spermatid nucleus; they now extend longitudinally over the whole length of the nucleus. Longitudinal sections of spermatids reveal as many as four or five rows of microtubules in longitudinal profile. The spermatids come to lie within deep recesses of the Sertoli cell cytoplasm (Fig. 14).

Enlarged detail of Fig. 5. We can notice the commencement of the acrosomal granule (AG). We also observed the subacrosomal space (SS) along with the Golgi apparatus (G) and the SER.
The acrosomal vesicle was taking the shape of the acrosomal cap by its internal changes. We can see the cytoplasm filled with an abundance of mitochondria (M), of Golgi bodies (G), and of SERs.
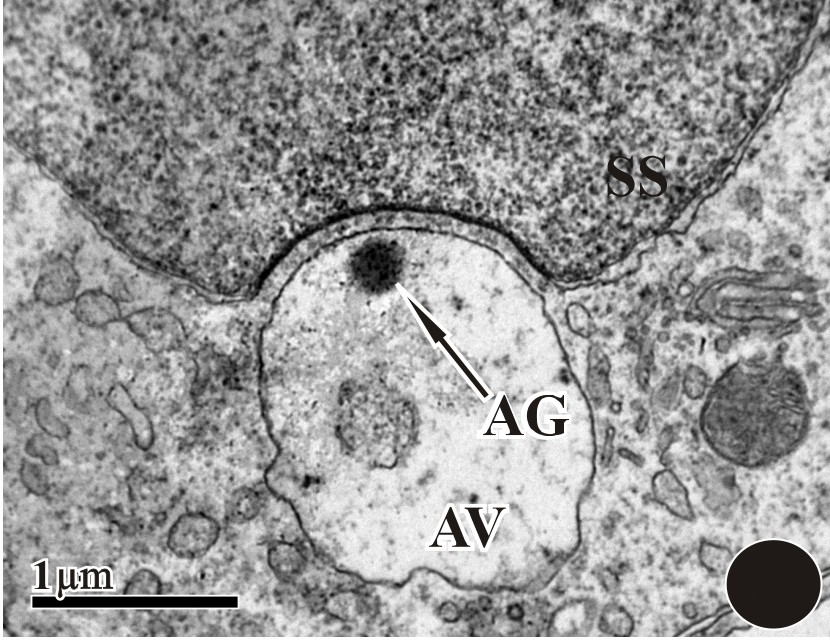
A large acrosomal vesicle; within it we can also find acrosomal granule (AG) and a small micro-vesicle. The subacrosomal space (SS) was distinct and filled with fluid.

Enlarged detail of Fig. 8, showing small vesicles moving towards acrosomal vesicles to fuse with them and discharge their contents inside the acrosome.

The nucleus (N) was engulfing an acrosomal vesicle. At the opposite side, we can see a budding tail region (arrow). The nucleus has become dark, with equally distributed chromatin.
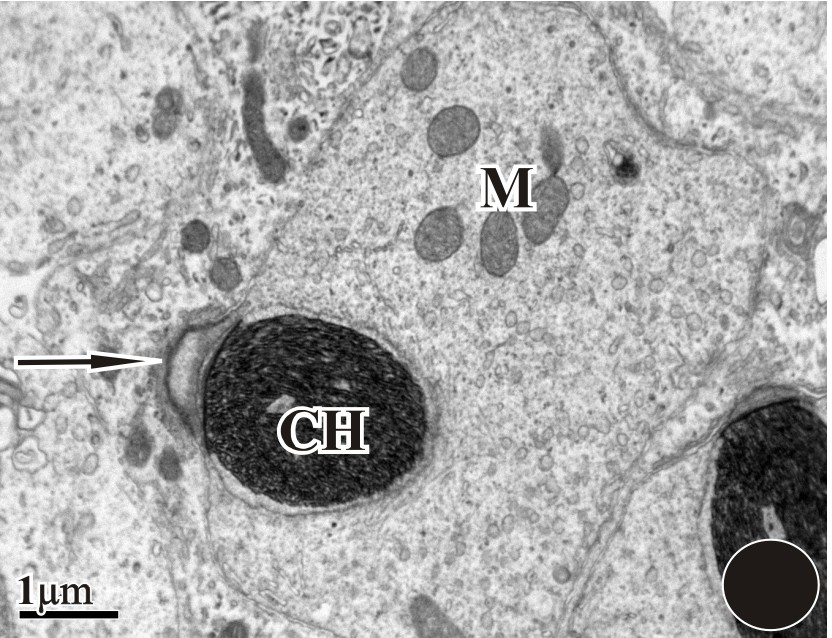
The transformation of round spermatid into elongated spermatid has taken place. An acrosomal vesicle can be seen in the anterior region.

A long nucleus with thread-like chromatin (CH) can be seen with a distinct acrosomal region. Many mitochondria were also seen in the vicinity.

A distinct acrosomal cap (AC) was present with subacrosomal space (SS). A large number of mitochondria (M) and SER were also present. Filamentous-like chromatin was seen in the nucleus. The nucleus was surrounded by a bilayer nuclear membrane.
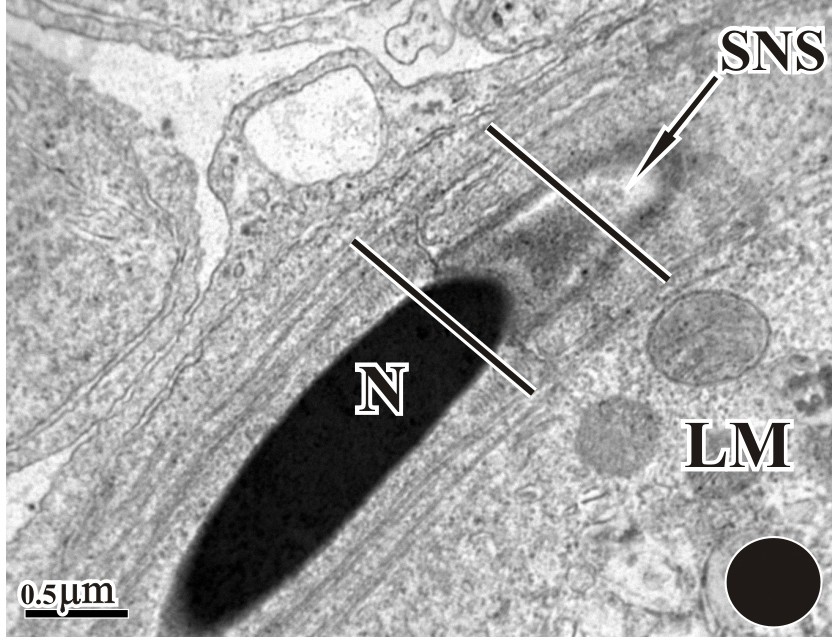
A nucleus with highly condense chromatin was present alongside with a subacrosomal nuclear space (SNC) and an acrosomal cap (AC). A longitudinal manchette was present, running parallel to the nucleus. Alongside the manchette, long microtubules can also be seen.
A small epinuclear lucent zone rests on the tip of the nuclear rostrum (Fig. 15) within the subascrosome space (Fig. 15). Within the nuclear rostrum of a mature spermatid, a distinct endonuclear canal exists and has an enclosed perforatorium (Fig. 16).

Endonuclear canal (nuclear gap) surrounded by a dark nucleus. The chromatin becomes more condensed in the nucleus (N) and forms large chromatin granules.
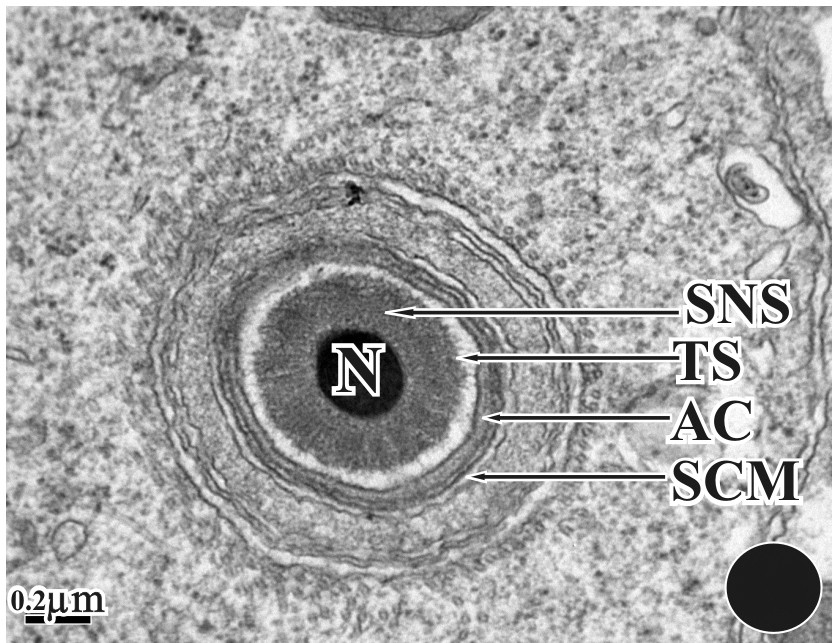
Cross-section of Fig. 14. Nucleus (N) surrounded by acrosome, subacrosomal nuclear membrane (SNC), transparent space (TS), microtubules and manchette. The inner and outer acrosomal membranes can also be seen.
4 Discussion
Most of the ultrastructure features of head differentiation within D. zarudnyi are similar to those described for other amphisbaenian reptiles. The early development of the acrosome complex within the D. zarudnyi testes is akin to what has been described for other reptiles. The single acrosome vesicle forms from transport vesicles delivered from the Golgi apparatus. This is comparable to what had been described for squamates [10,12,13] and other crocodilians [14,15]. In this study, we identified a unique feature in the early acrosome development in D. zarudnyi since a thin layer of endoplasmic reticulum forms a cap on the top of the developing acrosome vesicle. This process is not common in all lizards, except in a few species such as Alligator mississippiensis [16].
A single large acrosome granule forms and remains in contact with the inner acrosome membrane throughout sperm development until mid to late elongation when it diffuses uniformly throughout the acrosome lumen. The nucleus of the D. zarudnyi has uniform diffuse chromatin with small indices of heterochromatin, which is much different from the intermediate to heavily heterochromatic nuclei of Sphenodon [17] and chelonians [18]. Conversely, like in other lizards [14,15], the condensation of DNA in D. zarudnyi packs into large granules until the nucleus contains only homogeneous dark staining DNA, resembling the case of Sphenodon [17] and of chelonians [7,18]. In squamates, such as Agkistrodon piscivorus [13] and Anolis carolinensis [10], the DNA condenses in a filamentous helical fashion. In D. zarudnyi, early to mid-elongating spermatids have an adjacent endoplasmic reticulum that surrounds the nucleus, which may be an autoapomorphic feature of spermiogenesis in D. zarudnyi as this has not been reported in any other lizards including crocodilians [15]. The subacrosome space develops early in the round spermatid stage in D. zarudnyi and continues to enlarge during elongation, and this space accumulates a thick layer of dark staining granules similar to that described in other reptilian sauropsids [12,13]; once the acrosome has completed its development and growth, the nuclei of the early elongating spermatids become associated with the cell membrane. This contact with the cell membrane flattens the acrosome vesicle on the surface of the anterior nucleus, which aids in the migration of the acrosomal shoulders laterally over the apical nuclear head.
In chelonians [6,7,18] and other crocodilians [15], at least one endonuclear canal is present and houses the perforatorium, which extend deep into the nuclear body in middle-elongating spermatids. There has been some difference on whether a perforatorium exists within the endonuclear canals of some reptilian sauropsids [14]; however, in D. zarudnyi, a clearly visible perforatorium (dark stained) develops within the endonuclear canal, much like that observed in other lizards [14]. In contrast, all squamates studied to date have an extranuclear perforatorium (with no endonuclear canals) located in the subacrosome space within their spermatids and spermatozoa [10,12,13].
In D. zarudnyi, we observed a prominent longitudinal manchette associated with the nuclei of late elongating spermatids. We also noticed distinguished circular microtubules running parallel to the entire nucleus. The manchette has been implicated in the elongation process of the nucleus during spermiogenesis [19] and is considered a common structure observed during spermiogenesis in reptilian sauropsids [20]. There is only a single type of microtubule that makes up the circular and longitudinal manchette in D. zarudnyi, which is different from the case of the Caiman [14], where the microtubules of the longitudinal manchette are thicker-walled than those found in the circular manchette.
Although the characteristics of head differentiation within D. zarudnyi are similar to what has been described in other reptilians, there are three ultrastructural features of the D. zarudnyi spermatids that seem to be unique for this species:
- • the endoplasmic reticulum cap of acrosome development;
- • the concentric ring of ER around the nucleus of early- to middle-elongating spermatids;
- • the compound mitochondria of the midpiece that have cristae and concentric layers of cortical cristae.
D. zarudnyi spermatids also share one major feature with Sphenodon, chelonians, and other archosaurs, i.e. its endonuclear canals. These characteristics have not been reported for major reptilians. The importance of such differences and similarities in the structure of the reptilian spermatids between closely and distantly related species is unknown as there are not enough representative data from other reptilians. Until further morphological data on spermiogenesis and spermatozoa within reptilians are produced, answers to these questions will remain primitive. The ultrastructural study on D. zarudnyi head differentiation presented here provides a model that should be utilized in the future for evolutionary and phylogenic investigations.
Disclosure of interest
The authors declare that they have no conflicts of interest concerning this article.
Acknowledgments
The authors would like to sincerely acknowledge the deanship of Scientific Research at King Saud University for having funded this research through the Research Group Project No. RGP-VPP-289.