

1 Introduction
In recent publications, we have already outlined a number of difficulties with the taxonomy of the subfamily Scorpiopinae [1,2]. Morphological assessments demonstrated that the subfamily Scorpiopinae forms a monophyletic group within Euscorpiidae and does not share any synapomorphies with North American Vaejovidae [3]. The tribe Scorpiopini includes six Asian genera, mainly from the southern and southeastern regions of the continent. The composition of these genera is largely due to Vachon [4], who revised Scorpiops and described three new subgenera – Alloscorpiops, Euscorpiops and Neoscorpiops – in addition to the nominotypical subgenus, Scorpiops. These four subgenera were later elevated to generic rank by Lourenço [5], who added the monotypic genera Parascorpiops Banks 1928 and Dasyscorpiops Vachon, 1974 to the tribe, bringing the total number of genera to six. Recently, Lourenço [6] divided the genus Alloscorpiops into two subgenera with the creation of the subgenus Laoscorpiops. This decision was rejected by Kovařík et al. [7], based on unreliable arguments that attributed undue importance to the homologies defined by Vachon [8]. In fact, the study carried by Vachon [8] was largely unfinished and Vachon himself (in litt.) admitted a number of difficulties relating to his definitions of homologies. For these reasons, we prefer to maintain the validity of the subgenus Laoscorpiops stat. n. until further studies on the trichobothrial patterns are carried out. Other aspects concerning a number of taxonomic decisions have been summarized in previous papers [1,2,6,9] and will not be further discussed at present. Here, we describe a new subgenus and species belonging to the genus Scorpiops, collected in a limestone formation covered by rainforest, in the extreme North of Vietnam. This new scorpion taxon may represent yet another endemic element for the fauna of Southeast Asia, mainly Vietnam and Laos.
2 Methods
The new scorpion was found on the wall of a limestone formation, in a rainforest zone. Illustrations and measurements were made using a Wild M5 stereomicroscope with a drawing tube and an ocular micrometer. Measurements follow Stahnke [10] and are given in millimetres. Trichobothrial notations follow Vachon [8] and morphological terminology mostly follows Vachon [11] and Hjelle [12].
3 Taxonomic treatment
Family EUSCORPIIDAE Laurie, 1896
Subfamily SCORPIOPINAE Kraepelin, 1905
Genus Scorpiops Peters, 1861
Subgenus Vietscorpiops subgen. n.
Etymology: the subgeneric name associates Scorpiops with Vietnam, the country where the new subgenus was found
Diagnosis of the new subgenus
The new subgenus presents most of the characteristics already defined for the genus Scorpiops [3,4]. It can, however, be characterized by a number of distinct features: two lateral eyes represented by lenses; a possible third pair is represented by inconspicuous maculae without lenses. An internal apophysis is present on the movable chelal finger; this apophysis lodges in a depressed zone of the fixed finger when the chela is closed, then making it impossible to see from the external aspect (Figs. 1 and 3). This character was not mentioned by either Vachon [4] or Soleglad and Sissom [3]. Trichobothrium Dt of chela situated at same level as Trichobothrium Eb3. Telson annular ring strongly marked (see next section).

(Color online.) Scorpiops (Vietscorpiops) dentidactylus sp. n., male holotype. A. Anterior edge of carapace, showing two pairs of lateral eyes. B. Internal aspect of chela, showing the conspicuous apophysis on the movable finger.
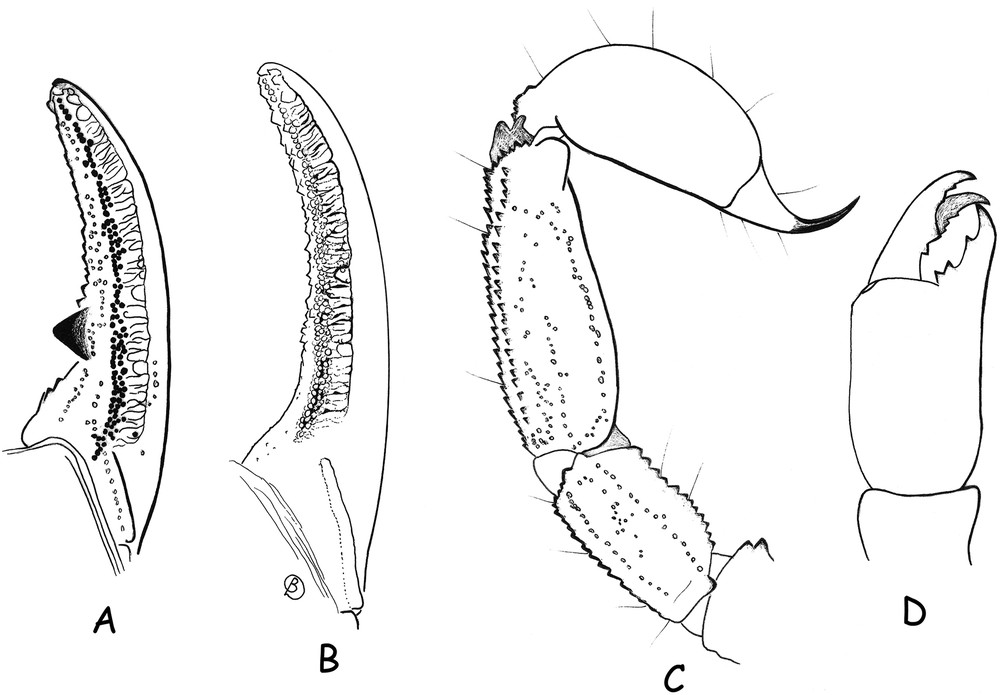
Scorpiops (Vietscorpiops) dentidactylus sp. n., male holotype (A, C and D). Scorpiops (Scorpiops) dakrong, male holotype (B). A and B. Cutting edge of chelal movable fingers, respectively showing the presence and absence of a conspicuous apophysis. C. Metasomal segment V and telson, lateral aspect. D. Chelicera, dorsal aspect.
Type species: Scorpiops (Vietscorpiops) dentidactylus sp. n.
The taxonomic positions of the genera Scorpiops and Euscorpiops
Vachon [4] and Soleglad and Sissom [3] supported the validity of both Scorpiops and Euscorpiops based on the position of chelal trichobothrium Eb3 and the presence or absence of an annular ring on the telson. This dichotomy, however, is not always evident in all studied species. A first exception was noted by Lourenço and Pham [2] in S. oligotrichus Fage. Although the position of chelal trichobothrium Eb3 (basal or at the same level of Dt) associates this species with the genus Scorpiops, the presence of a weakly to moderately marked annular ring on the telson is a feature shared with Euscorpiops. A second example is the recently described species Euscorpiops dakrong Lourenço and Pham, also from Vietnam [2]. In this case, trichobothrium Eb3 is basal or at the same level as Dt, but the annular ring on the telson is strongly marked. Faced with these complicated cases, we consider it necessary to give priority to the position of trichobothrium Eb3 in relation to Dt for generic assignment of the species. It seems that the presence of an annular ring on the telson is a more general character than previous believed. Consequently, S. oligotrichus is maintained in the genus Scorpiops, whereas Euscorpiops dakrong is transferred to the genus Scorpiops as S. dakrong (Lourenço and Pham, 2014) n. comb.
Scorpiops (Vietscorpiops) dentidactylus sp. n. (Fig. 2)

(Color online.) Scorpiops (Vietscorpiops) dentidactylus sp. n. A and B. Male holotype. Habitus, dorsal and ventral aspects (scale bars = 5 mm).
Type material. Vietnam, Dien Bien Province, Muong Nhe District, Nam Vi Commune, rainforest, limestone formation (C. Dawidoff leg.) 1938–1939. Male holotype deposited in the Muséum national d’histoire naturelle, Paris.
Etymology: The specific name refers to the large “tooth” found on the movable chelal finger.
Diagnosis: As for the subgenus.
Description: Total body length medium to small, male 30.6 mm. Body and pedipalps moderately slender. Coloration reddish-yellow to reddish-brown. Carapace reddish-brown with paler zones on posterior edge and on furrows. Tergites reddish-yellow, paler than carapace. Metasomal segments reddish-yellow; telson dark yellow; base of aculeus yellow and tip reddish. Chelicerae yellow with diffuse, variegated spots, these being better marked at the base of the teeth. Pedipalps reddish-brown; fingers darker than chela manus. Legs dark yellow. Venter yellow, with a few infuscations; sternite VII darker than the other sternites.
Morphology. Carapace moderately to strongly granular; furrows moderately deep. Median eyes anterior to middle of carapace; two pairs of lateral eyes with lenses; the third pair only represented by inconspicuous maculae, without lens. Sternum pentagonal, wider than long. Tergites moderately granulated; VII with five carinae, moderately to strongly marked. Pectines very large; pectinal tooth count 7-7; fulcra vestigial. Sternites smooth and punctate; spiracles oval in shape; sternite VII with four weakly marked carinae and vestigial granulation. Metasomal segment I wider than long; segment II as long as wide; III to V longer than wide; 10-10-8-8-7 carinae present on segments I–V; intermediate carina on segment II represented by a few posterior granules; dorsal carinae on segments II–IV with posterior spinoid granules; metasomal tegument moderately to strongly granulated; ventral carina on segment V with strongly marked spinoid granules. Telson vesicle smooth, without granulation. Setation moderate on metasomal segments and telson. Pedipalps: femur with dorsal internal, dorsal external, ventral internal and ventral external carinae moderately to strongly marked; tegument moderately granular. Patella with dorsal internal, dorsal external, ventral internal, ventral external and external carinae moderately to strongly marked; two moderately marked spinoid granules of similar size present on internal aspect; tegument moderately granular. Chela with dorsal marginal, external secondary, ventral internal and ventral carinae moderately marked; other carinae moderately to weakly marked; tegument weakly granulated dorsally and strongly granulated ventrally. Chelal fingers with two longitudinal series of granules almost fused; inner and outer accessory granules present. Internal face of movable chelal finger with a strong and peculiar apophysis (see taxonomic comments). Cheliceral dentition as defined by Vachon (1963, 1980[4,13]); movable finger with reduced teeth and 4–5 teeth on ventro-internal face (Fig. 3). Trichobothriotaxy of type C, as shown in Fig. 4 [8]. Trichobothrial pattern with three trichobothria on femur: dorsal, internal and external. Patella with 2 dorsal, 1 internal, 9 ventral and 17 external trichobothria. Chelal manus with 4 ventral, 2 dorsal (Dt, Db), 2 internal (ib, it), 1 Est, 5 Et, 1 Esb and 3 trichobothria in the Eb series. Trichobothrium Dt of chela is situated at the same level as Trichobothrium Eb3 [3,4].
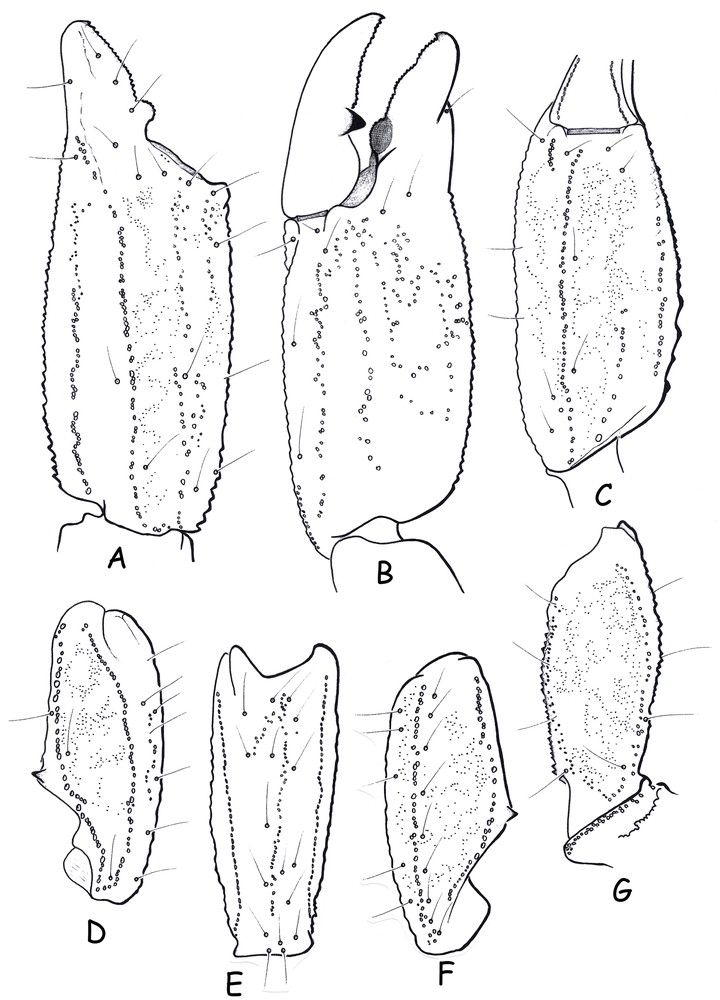
Scorpiops (Vietscorpiops) dentidactylus sp. n., male holotype. Trichobothrial pattern. A–C. Chela, dorso-external (A), internal (B) and ventral (C) aspects. D–F. Patella, dorsal (D), external (E) and ventral (F) aspects. G. Femur, dorsal aspect.
Morphometric values (in mm) of male holotype. Total length (including telson) 30.6. Carapace: length 4.6; anterior width 2.8; posterior width 4.4. Mesosoma: length 9.5. Metasomal segment I: length 1.6, width 2.2; II: length 2.0, width 2.0; III: length 2.1, width 1.9; IV: length 2.4, width 1.8; V: length 4.1, width 1.8, depth 1.5. Telson: length 4.3. Vesicle: width 1.8, depth 1.6. Pedipalp: femur length 4.4, width 2.0; patella length 4.8, width 1.9; chela length 8.3, width 2.7, depth 2.3; movable finger length 3.4.
4 Taxonomic comments on two characters defining the new subgenus
The number of lateral eyes is one of the characters used to define genera and even, in some cases, families. In the subfamily Scorpiopinae, only the genus Parascorpiops Banks is defined as having two pairs of lateral eyes; all the other known genera present three pairs. In the case of Scorpiops (Vietscorpiops) dentidactylus subgen. n. et sp. n., only two pairs of lateral eyes with lenses are observable (Fig. 1); a possible third pair is represented by inconspicuous maculae, but these lack lenses. The study of further specimens of the new species will be necessary to determine whether this character is stable within the population.
The presence of an internal apophysis on the movable chelal finger, which lodges in a depressed zone of the fixed finger when the chela is closed (Figs. 1 and 3) appears to be unique among Scorpiopinae, not having been recorded previously in this group of scorpions [3,4]. A precise analysis of this character shows that it has no similarities with the classical scalloping structure observed on the proximal dentate margin of the fixed chelal finger observed in species of several other scorpion families, in which it is usually more pronounced in males than in females. The function of such structures in these groups is primarily associated with the apprehension capacities of chelal fingers. However, the internal apophysis on the movable chelal finger of Scorpiops (Vietscorpiops) dentidactylus subgen. n. et sp. n. instead shows affinities with structures observed in other scorpion families and associated with secondary sexual characters (e.g., the genus Bothriurus Peters, family Bothriuridae). These structures, such as the apophysis present on the internal face of the chela hand of male specimens of Bothriurus, play an important role during sexual courtship and mating [14]. It can be suggested that the apophysis observed on the movable chelal finger of Scorpiops (Vietscorpiops) dentidactylus subgen. n. et sp. n., plays a similar role in the biology of this species.
5 Ecological aspects of Muong Nhe Nature Reserve
Muong Nhe Nature Reserve, Nam Vi commune, Muong Nhe District, Dien Bien Province is located in the extreme northwest region within the tri-border area of Vietnam, Laos and China (Fig. 5). The reserve is contiguous with forest in Laos but separate from forested areas in China due to clearance for agriculture on the Chinese side of the border. The terrain of the reserve is dominated by medium-high mountains and covered by evergreen forest, however many areas in the border area of Vietnam and Laos are covered by grassland.

Map of Vietnam showing the type locality of Scorpiops (Vietscorpiops) dentidactylus sp. n. (black circle).
The typical landscape and terrain of the nature reserve consists of medium and high mountains. There is one main mountain range, which runs along the Laos/Vietnam border called Phu Den Dinh with the highest peak named Pu Pá Kun (1892 m above sea level). The Ta Long San, Phu Tu Na ranges in the border area with China reach 1405 m. The average height of the mountains in the reserve is lower than 1000 m. Within the ranges are narrow valleys and streams running from northwest to southeast.
Weather in the area represents the typical inland tropical climate of the northwest mountainous provinces. The area is protected by the Hoang Lien Son ranges and therefore it is little affected by the Siberian north wind. There are two typical seasons in the area, the rainy season with high temperatures and humidity from April to October and the dry season with cold and dry weather from November to April. The average annual temperature is 22.5 °C with highest temperatures reaching 39 °C and lowest reaching 7 °C. The average annual rainfall is 1950 mm and is focused mostly in summer (June to August) and the average annual humidity is 85%.
Preliminary botanical surveys in the reserve have confirmed that there are 740 species of vascular plant, which belong to 500 genera of 156 families and five orders of plant. Many of the plant species are listed as endemic and endangered species in the Vietnam Red Data Book. Three broad forest types have been observed:
- a. Tropical monsoon forest – this type of forest is distributed in areas lower than 800 m asl, mostly on sloping areas of the mountain ranges. These forests are mostly impacted by human activities, such as selective logging and shifting cultivation. The new subgenus and species was found in this type of formation (Fig. 6);
- b. Semi-tropical monsoon forest – this type of forest is distributed between elevations of 800 to 1800 m asl and is found mostly in medium and high mountain areas. This forest type is mostly intact and is distributed on highly sloped sides of the mountain ranges in Muong Nhe;
- c. Grassland formed after shifting cultivation and forest fire – this type of vegetation is found mostly in areas close to local residential areas where shifting cultivation is still the main agricultural practice. Many areas are now cover by tall grassland and bush, as they have been abandoned by local people, and are currently included within the nature reserve's boundaries. Areas close to the international Laos/Vietnam border are now mostly covered by grassland formed by annual forest fires. Information from the management board suggests that forest fires are created by humans, mostly hunters during the dry season, to make new grasslands for hunting [15,16].

(Color online.) Typical evergreen lowland rain forest vegetation of Muong Nhe Nature Reserve, type locality of Scorpiops (Vietscorpiops) dentidactylus sp. n.
Disclosure of interest
The authors declare that they have no conflicts of interest concerning this article.
Acknowledgements
We are very grateful to Élise-Anne Leguin (Muséum, Paris) for her contribution to the preparation of the photographs and plates, to Mark Judson (Muséum, Paris) for revising an earlier version of the text, and to two anonymous referees for some useful comments to the text.