

1 Introduction
The gastrointestinal (GI) tract is supplied by different types of neurons. The extrinsic innervation of the esophagus, the stomach, and the intestine comprises projections of parasympathetic [1], sympathetic [2], and spinal neurons [3]. Parasympathetic efferent innervation of the major part of the digestive tract (from the esophagus to the transverse colon) is supplied by the vagal nerve and neurons located within dorsal motor nucleus of the vagal nerve (DMV) within the brainstem [4,5]. However, the posterior segments of the GI tract (from the descending colon to the anus) are innervated mainly by projections of neurons positioned in dorsal intermediolateral column of the sacral spinal cord (neuromeres S1–S4) [4,6]. Sympathetic efferent innervation of the digestive tract is realized by postganglionic nerves, which are the projections of neuronal cells located in the sympathetic chain ganglia and prevertebral ganglia of the abdominal and pelvic areas, such as the celiac, superior, and inferior mesenteric or pelvic ganglia [2,7]. The exact distribution of these neurons clearly depends on the innervated by them fragments of the digestive tract. The third component of the extrinsic innervation of the digestive tract is an afferent innervation, which conveys sensory and pain stimuli from the GI tract to the central nervous system. Sensory neurons supplying the digestive tract are located in the nodose ganglia of the vagal nerve [8] or the dorsal root ganglia [3,9] in various neuromeres (according to the innervated fragment of the GI tract).
Apart from the extrinsic innervation, the GI tract is supplied by the enteric nervous system (ENS), located in the wall of the digestive tract and containing millions of neuronal cells grouped in ganglionated plexuses [10–13]. The types of these plexuses depend on the animal species and the fragment of the digestive tract. In large domestic mammals (including the pig), the ENS in the esophagus and stomach is built of two types of plexuses: the myenteric plexus–located between longitudinal and circular muscle layers and submucous plexus–near the lamina propria of the mucosal layer, whereas, in the intestine, the submucous plexus is divided into outer submucous plexus–positioned along the inside of the circular muscle layer and inner submucous plexus–situated like the submucous plexus in rodents [14]. Enteric neurons show wide variations in terms of their morphology, functions, and electrophysiological properties [10,15].
The most important criterion useful in the division of enteric neurons into various subclasses is their neurochemical coding [15]. Apart from acetylcholine–the classic neuromediator of the ENS, several dozens of other neuronal active substances acting as neuromediators and/or neuromodulators have been described in enteric neuronal structures [16,17]. The most important of them are: vasoactive intestinal polypeptide (VIP), nitric oxide (NO), galanin (GAL), substance P (SP), and many other ones [16,18,19]. From a wide range of neuronal factors, that have been described in enteric nervous structures, cocaine- and amphetamine-regulated transcript peptide (CART) is one of the less known substances. Till now CART has been described in various parts of both central and peripheral nervous systems, especially within the ENS [20,21], where this peptide has been noted in the myenteric and submucous enteric plexuses and muscular and in mucosal intestinal layers of various mammal species, including humans, and the number of them clearly depended on the fragment of the digestive tract studied [21–23].
Contrary to the central nervous system, where first of all CART is known as a main anorectic peptide involved in the regulation of food intake [21], exact roles of this peptide in the ENS still remain unclear. Admittedly previous studies showed that CART may reduce gastric acid secretion and stimulate colonic motility [21]. The mechanisms of these actions, which most likely proceed with the participation of the central nervous system [20,24] are unknown. Moreover, especially high expression of CART in myenteric plexuses and intramuscular nerve fibers [25,26] can suggest the important role of this peptide in intestinal motility, and changes in its expression under various pathological processes [14,27] denote neuroprotective and/or neurotrophic activities.
Contrary to the stomach and intestine, the knowledge concerning the distribution and functions of CART in the esophageal ENS is very limited [23]. Therefore, the aim of the present investigation was the exact determination of the distribution of CART, as well as the colocalization of this peptide with other better known neuronal factors in the ENS of the esophagus in the domestic pig. It should be underlined that this species, due to considerable similarities to human with respect to the anatomy and physiology of enteric neurons, is often used as an animal model for studying the processes taking place in the human digestive tract [28].
2 Materials and methods
The present experiment was carried out using six immature female pigs of the Large White Polish breed (approximately 8 weeks old and 18 kg of body weight). During the investigation, pigs were kept under standard laboratory conditions. The animals were housed in pens with an area of about 4 m2 (three pigs in one pen) with unlimited access to water and nourished twice a day with complete feed appropriate to species and age. All procedures connected to the experiment were performed according to the instructions of the Local Ethical Committee, Olsztyn (Poland).
After a four-day adaptation period, pigs were pre-treated with Stressnil (Janssen, Belgium, 75 μl/kg of body weight, i.m.) and, after 15 min, were subjected to euthanasia by an overdose of sodium thiopental (Thiopental, Sandoz, Kundl, Austria) given intravenously. Then animals were perfused transcardially with 4% buffered paraformaldehyde prepared ex tempore. Two-centimeter-long segments of the cervical, thoracic, and abdominal esophagus were collected. They were post-fixed by immersion in 4% buffered paraformaldehyde for 30 min, rinsed in phosphate buffer for three days, and stored at 4 °C in 18% buffered sucrose solution for at least two weeks. Then samples were frozen at −25 °C and cut into 10-μm-thick sections using a microtome (Microm, HM 525, Walldorf, Germany).
Then the fragments of esophagus were subjected to standard double- and triple-labeling immunofluorescence techniques. After removal from the freezer, glasses with fragments of esophagus were dried at room temperature (rt) for 45 min and rinsed in PBS (pH 7.4, three times for 15 min). Then they were incubated with a blocking solution containing 10% normal goat serum, 0.1% bovine serum albumin, 0.01% NaN3, Triton x-100, and thimerozal in PBS for 1 h (rt). After a renewed rinse in PBS (3 × 10 min), tissues were incubated (overnight; rt, in a humid chamber) with a mixture of two (in the case of double immunofluorescence technique) or three (in the case of triple immunofluorescence technique) antibodies obtained from different species and directed towards CART and other one (double immunofluorescence technique) or two (triple immunofluorescence technique) selected neuronal substances, i.e. protein gene product 9.5 (PGP 9.5, used here as pan-neuronal marker), vesicular acetylocholine transporter (VAChT, used here as marker of cholinergic neurons), neuropeptide Y (NPY), vasoactive intestinal polypeptide (VIP), somatostatin (SOM), galanin (GAL), substance P (SP), calcitonin gene related peptide (CGRP), leu-enkephalin (LENK) or neuronal isoform of nitric oxide synthase (nNOS, a marker of nitrergic structures). The precise specification of antisera is presented in Table 1. The next day, the tissues were again rinsed in PBS (3 × 10 min), and complexes of primary antibodies bound to appropriate antigens were visualized by incubation (1 h, rt) with species-specific secondary antisera conjugated to FITC or biotin. Then fragments of esophagus were rinsed in PBS (3 × 15 min) and the secondary antibodies conjugated to biotin were visualized by incubation of tissues with a streptavidin–CY3 complex (1 h, rt). After a renewed rinse in PBS (3 × 10 min), tissues were flooded with buffered glycerol and covered with the cover slip. The esophagus fragments prepared in this way were stored at 4 °C until microscopic examinations. Standard controls, i.e. pre-absorption of the neuropeptide antisera with appropriate antigen, omission, and replacement of primary antisera by non-immune sera were performed to test the antibodies and the specificity of the method.
List of antisera and reagents used in immunohistochemical investigations.
| Primary antibodies | ||||
| Antisera | Code | Host Species | Dilution | Supplier |
| PGP 9.5 | 7863-2004 | Mouse | 1:2000 | Biogenesis, Poole, UK |
| CART | H-003-61 | Rabbit | 1:20,000 | Phoenix, Aachen, Germany |
| GAL | T-5036 | Guinea Pig | 1:1000 | Peninsula, San Carlos, CA, USA |
| CGRP | T-5027 | Guinea Pig | 1:1000 | Peninsula |
| nNOS | N2280 | Mouse | 1:2000 | Sigma, St Louis, MO, USA |
| NPY | NZ1115 | Rat | 1:300 | Biomol Research Laboratories Inc, Plymouth, PA, USA |
| SOM | 8330-0009 | Rat | 1:100 | Biogenesis |
| VAChT | H-V007 | Goat | 1:2000 | Phoenix |
| VIP | 9535-0504 | Mouse | 1:2000 | Biogenesis |
| LENK | LS-C123457 | Mouse | 1:1000 | LifeSpan Biosciences Inc, Seattle, WA, USA |
| SP | SP-DE4-21 | Mouse | 1:1000 | Abcam, Cambridge, UK |
| Secondary antibodies | ||||
| Reagent | Code | Dilution | Supplier | |
| FITC-conjugated donkey–anti-mouse IgG (H + L) | 715-095-151 | 1:800 | Jackson, West Grove, PA, USA | |
| FITC-conjugated donkey–anti-rat IgG (H + L) | 712-095-153 | 1:800 | Jackson | |
| FITC-conjugated donkey–anti-guinea pig IgG (H + L) | 706-095-148 | 1:1000 | Jackson | |
| FITC-conjugated donkey–anti-goat IgG (H + L) | 705-096-147 | 1:1000 | Jackson | |
| Biotinylated goat-anti-rabbit immunoglobulins | E 0432 | 1:1000 | DAKO, Utrecht, The Netherlands | |
| AMCA-conjugated donkey–anti-mouse IgG (H + L) | 715-155-151 | 1:50 | Jackson | |
| AMCA-conjugated donkey–anti-goat IgG (H + L) | 705-156-147 | 1:50 | Jackson | |
| CY3-conjugated streptavidin | 016-160-084 | 1:9000 | Jackson |
To determine the overall percentage of CART-positive neuronal cells, at least 500 cell bodies, in which a pan-neuronal marker PGP 9.5 was present, were examined for CART-like immunoreactivity in the myenteric (MP) and submucous plexus (SP) of the cervical, thoracic, and abdominal parts of the esophagus of each studied animal. In this case, the neurons immunoreactive to PGP 9.5 included into the study were treated as 100%. For example, if in the MP of the selected fragment of the esophagus of one animal, 567 PGP 9.5-LI neuronal cells (treated as 100%) would be counted and among them 263 would be immunoreactive to CART, the percentage of CART-positive neurons would amount to 46.38%. In turn, to determine the percentages of CART-positive neuronal immunoreactive to the particular other substances studied, at least 400 neurons immunoreactive to CART were evaluated for the presence of each neuronal factor included into the study (i.e. VAChT VIP, SOM, GAL, SP, CGRP, LENK or nNOS). In this part of the experiment, CART-positive perikarya were considered as representing 100%. For example, if in the MP of the selected part of the esophagus of one animal, 482 CART-LI neurons (treated as 100%) would be counted and among them 145 cells would be immunoreactive to GAL, the percentage of CART-positive neuronal cells simultaneously immunoreactive to GAL would amount to 30.08%. The counting of each population of enteric neurons (PGP 9.5/CART, CART/VAChT CART/VIP, CART/SOM, CART/GAL, CART/SP, CART/CGRP, CART/LENK and CART/nNOS) was made up of at least 10 sections of each part of the esophagus in each animal, and the final number of sections included in the study depended on the individual variations in the ENS structure between particular pigs.
Double-labeled perikarya (only neurons with clearly visible nucleus were included) were determined under an Olympus BX51 microscope equipped with epi-fluorescence and appropriate filter sets, pooled and presented as mean percentage ± SEM. To prevent double counting of the same neuronal cell bodies, the esophagus sections included in the investigation were located at least 100 μm apart. All pictures were captured by a digital camera connected to a PC. Statistical analysis was carried out with ANOVA test (Graphpad Prism v. 6.0; GraphPad Software Inc., San Diego, CA, USA). The differences were considered statistically significant at P ≤ 0.05.
3 Results
3.1 Distribution of CART-LI neurons in the porcine esophagus
During the present study, CART-positive enteric neurons were noted both in myenteric and submucous plexuses of all parts of the porcine esophagus (Fig. 1). These values depended on the fragment of esophagus studied, but in all regions and each type of enteric plexuses, they were above 45% of all neurons immunoreactive to PGP 9.5 (Table 2, Fig. 1). Within the myenteric plexus, the percentage of CART-LI neurons in relation to all neurons within the particular types of plexuses immunoreactive to pan-neuronal marker PGP 9.5 amounted to 46.5 ± 5.7% in the cervical (Fig. 1, 1a), 58.1 ± 5.0% in the thoracic (Fig. 1, 2a) and 55.7 ± 2.1% in the abdominal (Fig. 1, 3a) parts of the esophagus. Numerous populations of CART-LI neuronal cells were also noted in the submucous plexus. In the cervical esophagus (Fig. 1, 1b) the percentage of CART-positive neurons (in relation to all PGP 9.5-LI cells) was higher than the values observed in MP of this fragment of the esophagus and amounted to 57.5 ± 11.7%. In the submucous plexus of the thoracic fragment of esophagus (Fig. 1, 2b) the percentage of CART-LI neurons achieved was 45.2 ± 0.9% of all cells immunoreactive to PGP 9.5, and in the abdominal part of esophagus (Fig. 1, 3b) this value amounted to 49.8 ± 4.3%.
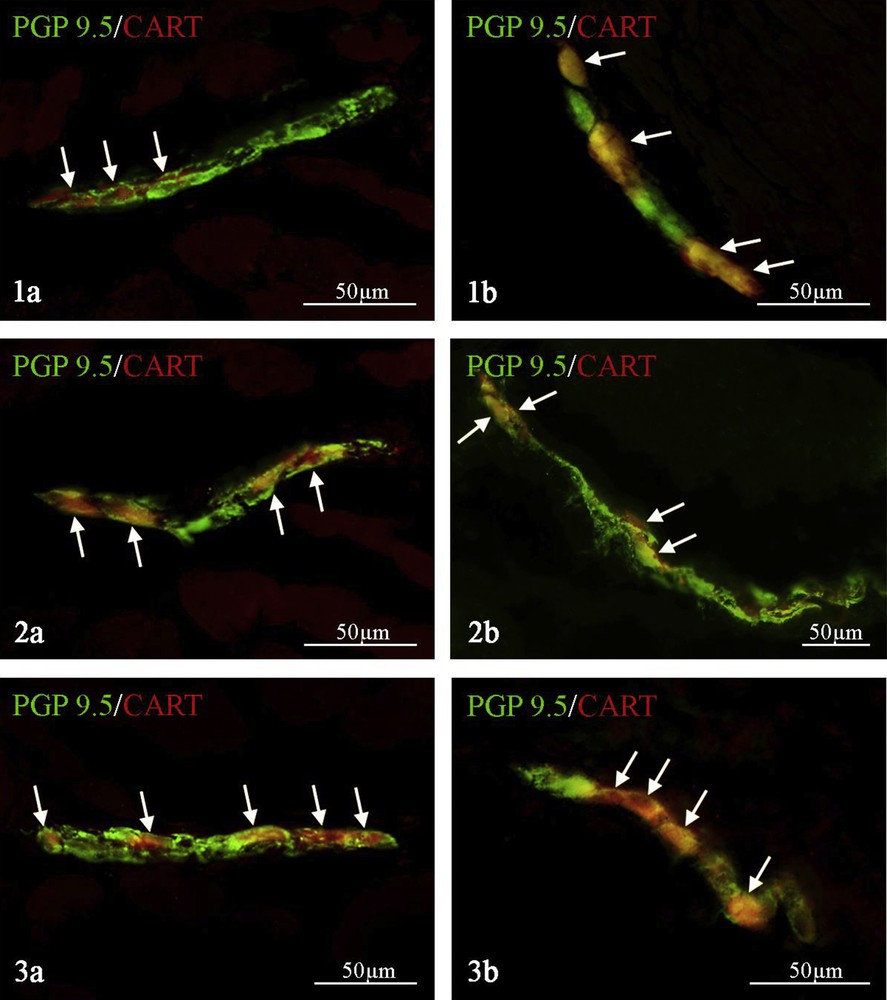
Neuronal cells immunoreactive to PGP 9.5–used as a panneronal marker (green) and CART (red) in the myenteric (a) and mucosal (b) plexuses of the cervical (1), thoracic (2), and abdominal (3) parts of the porcine esophagus. Images are composed of merged images taken separately from green and red fluorescent channels. PGP 9.5+/CART+ neurons are indicated with arrows.
Neurochemical characterization of CART-like immunoreactive (CART+) neurons in the enteric plexuses of the porcine esophagus. PGP 9.5 was used as a pan-neuronal marker that marks all neurons in particular types of plexuses, and so PGP 9.5+/CART+ cells illustrate the percentage of CART-positive neurons in relation to the all number of PGP 9.5–positive cells. In the case of co-localization of CART with other neurochemical factors, CART-positive neurons were considered as representing 100% for all combinations with other neuronal factors, and so all the values presented are percentages (means ± SEM) of CART+ neurons. Statistically significant data differences (P ≤ 0.05) in the percentages of the same neuronal population in the same types of enteric plexuses between particular fragments of the esophagus in particular animal groups are marked by different letters, and not significant data are marked by the same letters.
| Myenteric plexus (%) | Submucous plexus (%) | |
| Cervical esophagus | ||
| PGP 9.5+/CART+ | 46.5 ± 5.7a | 57.5 ± 11.7a |
| CART+/CGRP+ | 0 | 0 |
| CART+/GAL+ | 30.2 ± 10.0a | 29.1 ± 10.9a |
| CART+/LENK+ | 21.1 ± 4.9a | 9.7 ± 1.4a |
| CART+/nNOS+ | 70.0 ± 10.2a | 74.6 ± 5.4a |
| CART+/NPY+ | 17.0 ± 10.3a | 0a |
| CART+/SOM+ | 0 | 0 |
| CART+/SP+ | 0a | 0 |
| CART+/VAChT+ | 57.8 ± 6.9a | 53.5 ± 16.5a |
| CART+/VIP+ | 63.9 ± 0.9a | 100a |
| Thoracic esophagus | ||
| PGP 9.5+/CART+ | 58.1 ± 5.0b | 45.2 ± 0.9a |
| CART+/CGRP+ | 0 | 0 |
| CART+/GAL+ | 30.7 ± 3.3a | 100b |
| CART+/LENK+ | 19.5 ± 7.3a | 0b |
| CART+/nNOS+ | 57.4 ± 5.8b | 70.0 ± 4.2a |
| CART+/NPY+ | 19.6 ± 7.2a | 0a |
| CART+/SOM+ | 0 | 0 |
| CART+/SP+ | 6.0 ± 3.0b | 0 |
| CART+/VAChT+ | 86.9 ± 2.9b | 78.6 ± 14.8b |
| CART+/VIP+ | 72.4 ± 1.9b | 90.0 ± 10.0a |
| Abdominal esophagus | ||
| PGP 9.5+/CART+ | 55.7 ± 2.1a | 49.8 ± 4.3a |
| CART+/CGRP+ | 0 | 0 |
| CART+/GAL+ | 36.4 ± 5.2b | 60.0 ± 2.0c |
| CART+/LENK+ | 2.0 ± 1.1b | 10.3 ± 4.0a |
| CART+/nNOS+ | 80.0 ± 3.8c | 86.5 ± 4.5b |
| CART+/NPY+ | 56.2 ± 2.4b | 77.4 ± 12.6b |
| CART+/SOM+ | 0 | 0 |
| CART+/SP+ | 4.3 ± 4.1b | 0 |
| CART+/VAChT+ | 97.2 ± 1.6c | 82.1 ± 16.0b |
| CART+/VIP+ | 78.4 ± 4.0b | 83.6 ± 3.0b |
3.2 Colocalization of CART with other neuronal factors in enteric neurons of the porcine esophagus
The presence of a wide range of other neuronal factors was observed in CART-LI esophageal neurons during the present investigation. The degree of colocalization of CART with particular substances varied considerably between types of plexuses, as well as the esophagus fragment (Table 2).
3.2.1 Cervical part of esophagus
CART-LI neurons within the myenteric plexus of cervical esophagus (Fig. 2A) most often showed also the presence of NOS and/or VIP. The percentage of CART+/nNOS+ neurons in relation to all perikarya immunoreactive to CART amounted to 70.0 ± 10.2%, and in the case of CART+/VIP+ cells this value reached 63.9 ± 0.9% (Table 2). A slightly lower degree of colocalization of CART with VAChT and/or GAL was investigated (57.8 ± 6.9 and 30.2 ± 10.0% of all CART-LI neurons, respectively). In turn, neuronal cells that showed the simultaneous presence of CART and LENK or CART and NPY were less numerous. The percentage of such perikarya amounted to 21.1 ± 4.9% in the case of CART+/LENK+ neurons and 17.0 ± 10.3% for CART+/NPY+ cells (Table 2). However, the presence of other studied neuronal factors, such as CGRP, SOM, and SP, was not observed in CART-positive neuronal cells in the myenteric plexus of the cervical part of the esophagus (Table 2).
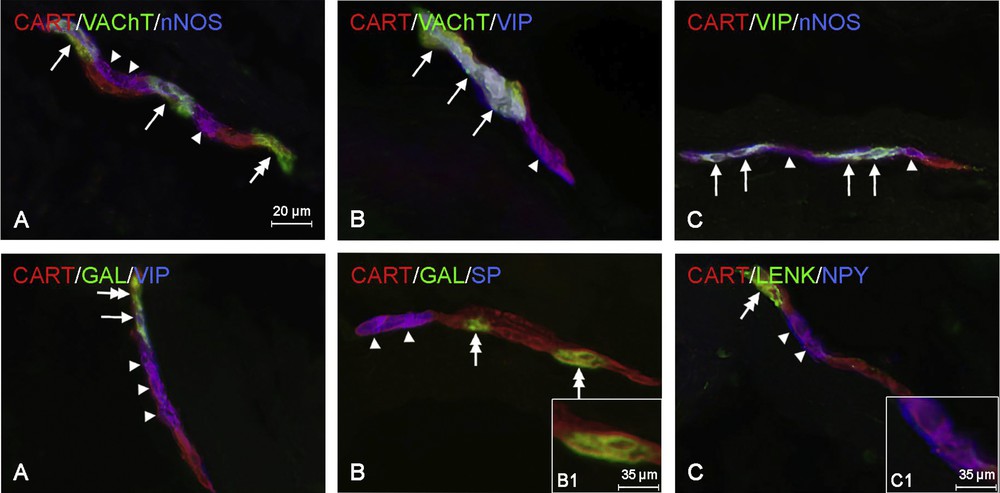
Representative images of co-localization of CART with other active substances in the myenteric plexus of the cervical (A), thoracic (B), and abdominal (C) parts of the porcine esophagus. Images are composited of merged images taken separately from blue, green, and red fluorescent channels. Top row: CART+/VAChT+/nNOS+ neurons are indicated with arrows (A), CART+/VAChT–/nNOS+ neurons are indicated with arrowheads, the CART+/VAChT+/nNOS– neuron is indicated with a double-headed arrow. B. CART+/VAChT+/VIP+ neurons are indicated with arrows, the CART+/VAChT–/VIP+ neuron is indicated with an arrowhead. C. CART+/VIP+/nNOS+ neurons are indicated with arrows, CART+/VIP–/nNOS+ are indicated with arrowheads. Bottom row: the CART+/GAL+/VIP+ neuron is indicated with an arrow (A), CART+/GAL–/VIP+ neurons are indicated with arrowheads, the CART+/GAL+/VIP– neuron is indicated with a double-headed arrow (B) CART+/GAL–/SP+ neurons are indicated with arrowheads, CART+/GAL+/SP–neurons are indicated with double-headed arrows. B1. CART+/GAL+/SP–neuron. C. CART+LENK–/NPY+ neurons are indicated with arrowheads, the CART+/LENK+/NPY– neuron is indicated with a double-headed arrow. C1. CART+/LENK–/NPY+ neuron.
In the submucous plexus of the cervical esophagus (Fig. 3A), all CART-positive neuronal cells showed the simultaneous presence of VIP. The degree of colocalization of CART and nNOS and/or VAChT was also high and similar to those seen in the myenteric plexus. Namely, 74.6 ± 5.4% neurons immunoreactive to CART showed the presence of NOS, and in the case of CART+/VAChT+ cells, this value amounted to 53.5 ± 16.5% (Table 2). A slightly lower number of CART-LI neurons (like in the MP) were also GAL–positive (29.1 ± 10.9%). In turn, the degree of colocalization of CART with LENK or NPY in the submucous plexus was considerably lower than those observed in the myenteric plexus (Table 2). Only 9.7 ± 1.4% of all neurons immunoreactive to CART showed the presence of LENK, and the colocalization of CART and NPY was not observed at all. Moreover, as in the myenteric plexus, the presence of CGRP, SOM, and SP in CART-positive neurons in the submucous plexus was not noted (Table 2).
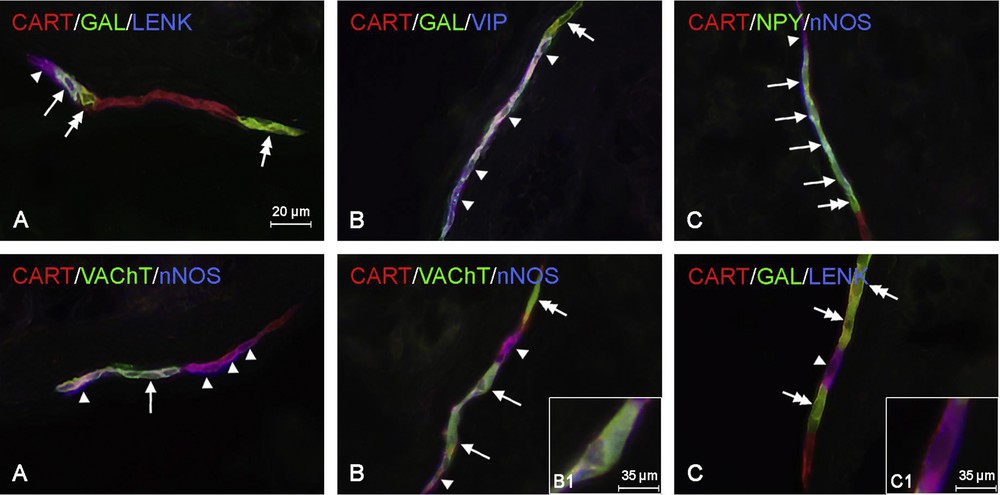
Representative images of the colocalization of CART with other active substances in the submucous plexus of the cervical (A), thoracic (B), and abdominal (C) parts of the porcine esophagus. Images are composed of merged images taken separately from blue, green, and red fluorescent channels. Top row: the CART+/GAL+/LENK+ neuron is indicated with an arrow, the CART+/GAL–/LENK+ neuron is indicated with an arrowhead, CART+/GAL+/LENK– neurons are indicated with double-headed arrows (A). B. CART+/GAL–/VIP+ neurons are indicated with arrowheads, the CART+/GAL+/VIP– neuron is indicated with a double-headed arrow. C. CART+/NPY+/nNOS+ are indicated with arrows, the CART+/NPY/nNOS+ neuron is indicated with an arrowhead, the CART+/NPY+/nNOS–neuron is indicated with a double-headed arrow. Bottom row: A. the CART+/VAChT+/LENK+ neuron is indicated with an arrow, CART+/VAChT–/nNOS+ neurons are indicated with arrowheads. B. CART+/VAChT+/nNOS+ neurons are indicated with arrows, CART+/VAChT–/nNOS+ are indicated with arrowheads, the CART+/VAChT+/nNOS– neuron is indicated with a double-headed arrow. B1. CART+/VAChT+/nNOS+ neuron. C. The CART+/GAL–/LENK+ neuron is indicated with an arrowhead, CART+/GAL+/LENK–neurons are indicated with double-headed arrows. C1. CART+/GAL–/LENK+ neuron.
3.2.2 Thoracic part of esophagus
In the thoracic part of the esophagus, the chemical coding of CART-positive cells was a little different from those observed in the cervical fragment. In the myenteric plexus (Fig. 2B), most CART-positive neurons were also immunoreactive to VAChT (86.9 ± 2.9% of all CART+ neurons) and/or VIP (72.4 ± 1.9%) (Table 2). In turn, the percentage of perikarya that were simultaneously immunoreactive to CART and nNOS was markedly lower than within the cervical esophagus and amounted to 57.4 ± 5.8%. Moreover, 6.0 ± 3.0% all CART-positive neurons showed the expression of SP, contrary to the cervical esophagus, where the colocalization of CART and SP was not observed. The degree of colocalization of CART with the other substances studied in the MP of the thoracic esophagus was similar to those observed in the cervical part of this organ and amounted to 30.7 ± 3.3%, 19.5 ± 7.3%, and 19.6 ± 7.2% in the case of CART+/GAL+, CART+/LENK+ and CART+/NPY+ neurons, respectively (Table 2).
Within the submucous plexus of the thoracic esophagus (Fig. 3B) all CART-positive perikarya were simultaneously immunoreactive to GAL (Table 2). The percentage of CART+/VIP+, CART+/VAChT+ and CART+/nNOS+ neurons was also high and amounted to 90.0 ± 10.0%, 78.6 ± 14.8%, 70.0 ± 4.2% of all neuronal cells immunoreactive to CART, respectively. The other studied substances were not observed in CART-LI perikarya within the submucous plexus of the thoracic esophagus (Table 2).
3.2.3 Abdominal part of the esophagus
In the myenteric plexus of the abdominal part of the esophagus (Fig. 2C), the vast majority of CART-positive neurons were also immunoreactive to VAChT (97.2 ± 1.6% of all CART+ perikarya), nNOS (80.0 ± 3.8%), and/or VIP (78.4 ± 4.0%) (Table 2). A slightly less large population of CART-LI neuronal cells showed simultaneously immunoreactivity to NPY (56.2 ± 2.4%) and/or GAL (36.4 ± 5.2%). In turn, SP and LENK were observed only within very small percentage of neurons immunoreactive to CART. These values amounted to 4.3 ± 4.1% and 2.0 ± 1.1%, respectively (Table 2). The presence of CGRP and SOM in CART-LI neuronal cells in the myenteric plexus was not observed, just like in the cervical and thoracic parts of the esophagus.
Within the submucous plexus (Fig. 3C), the majority of CART-positive neuronal cells showed immunoreactivity to nNOS (86.5 ± 4.5% of all CART+ cells), VIP (83.6 ± 3.0%) and/or VAChT (82.1 ± 16.0%), what is generally similar to situations in the other part of the esophagus (Table 2). A slightly lesser number of CART-LI neurons were also immunoreactive to NPY and/or to GAL. These values achieved 77.4 ± 12.6% and 60.0 ± 2.0%, respectively. In turn, the presence of LENK was noted in 10.3 ± 4.0% of all CART-positive neurons. The co localization of CART with other studied substances, such as CGRP, SOM and SP, was not observed in the submucous plexus within the abdominal part of the esophagus.
4 Discussion
During the present investigation, CART-immunoreactive neurons have been noted in both types of enteric plexuses within all studied fragments of the esophagus. Till now, the presence of this peptide has been noted in various parts of the peripheral nervous system, such as innervation of gall bladder, urinary bladder, pelvic ganglia, dorsal root ganglia, and many others [29–31]. The presence of CART has been also investigated in the GI tract of a wide range of mammal species, including humans [14,23,25]. It should be underlined that the number of CART-positive clearly depend on the mammal species and on the age of the studied animals, on the kind of fragment of the digestive tract and on the type of the enteric plexus [21]. In the light of the previous investigations on domestic pig, it is known that CART occurs in the neurons located in the myenteric plexus of the stomach, and it is not present in the enteric submucous ganglia [21,23]. Interestingly, the percentages of such neuronal cells are different in the reports of different authors. Namely, the percentage of CART-LI myenteric neurons in the gastric antrum under physiological conditions in pigs of about the same age fluctuated from about 40% [32] to above 50% [33] of all neurons, and in the gastric corpus–from about 14% to over 30% [32,33]. These differences suggest that the number of CART-positive neurons in the porcine ENS depends on not fully explained stimuli, such as minor changes in the feed and/or the presence of specific environmental microorganisms. Contrary to the stomach in the porcine intestine, CART has been observed in all types of the enteric plexuses. The percentage of CART-LI neurons in the small intestine (depending on the fragment studied) fluctuated from 10% to 20% of all neurons in the myenteric plexus, 3% to 10% in the outer submucous plexus and only to 2% in the inner submucous plexus [34]. In the large intestine, these values were as follows: from 2% to 5% in the myenteric plexus, about 5% in the outer submucous plexus and about 3% in the inner submucous plexus [35]. It should be pointed out that comparatively less is known about the distribution of CART-positive neurons within the esophagus. Till now, only one study on the subject has been published, where the presence of CART has been generally described in the muscular and mucosal plexuses of the human esophagus [23]. Relatively large number of CART-LI neurons (in relation to other parts of the GI tract described in the previous studies) noted during the present investigation in the porcine esophagus may suggest that the described peptide plays an important role in regulatory processes within this part of the digestive tract. In turn, similarities with the distribution of CART in the human [23] and porcine (this study) esophagi may confirm that resemblances in the organization of the ENS between human and domestic pigs [28] also concern the described fragment of the digestive tract.
It should be pointed out that, contrary to the central nervous system, where CART is known as an important factor taking part in the regulation of food intake, the exact functions of CART in the peripheral nervous system, including the innervation of the digestive system, still remain fragmentary and not fully elucidated. Previous studies showed that the described peptide may have an inhibitory effect on gastric acid secretion [20]. Moreover, it influences gastric and intestinal smooth muscles, and the character of this activity depends on the digestive tract fragment. In the stomach, CART shows relaxatory effects and inhibits gastric emptying, and, within the colon, stimulates intestinal motility [24]. However, due to the fact that direct administration of CART to intestinal cell cultures does not affect their function, the mechanisms of the above-mentioned actions remain pure conjecture, and are probably regulated via central nervous system [20,24].
The next important, but not fully explained aspect of CART activity in the digestive tract, is the participation of this peptide in pathological processes taking place in the stomach and the intestine. Previous studies described changes in CART expression in the enteric nervous structures under various pathological and toxicological factors [14,27], which can suggest participation of this substance in adaptive and/or neuroprotective processes within the ENS. This is in agreement with the previous studies on other parts of the nervous system, which showed that CART may be involved in neuronal development, neurotrophic, and neuroprotective effects, as well as regenerative mechanisms within the nervous tissue [21,29].
One of the ways to know the exact functions of CART in the ENS is the study on colocalization of this peptide with other better known neuronal factors. It is commonly known that one neuronal cell may produce even several active substances, which show similar activity [36,37]. The present study has shown that CART in the enteric neurons of the porcine esophagus colocalizes with a wide range of other neuronal factors.
In the light of the present investigation, CART in esophageal neurons colocalizes mainly with VAChT, VIP, nNOS, and GAL. Previous studies have also described a wide variety of CART-LI neurons in term of their neurochemical characterization in other parts of the digestive tract. It is known that CART in the enteric neuronal cells mainly co-localizes with choline acetyltransferase, GAL, substance P and nNOS, but the highest degree of co-localization has been described in the case of CART and VIP, because the level of coexistence of these substances in the same neuronal cells and nerves may achieve even 90% of all CART-LI structures [21,38]. Moreover, also other neuronal factors, including somatostatin, neuropeptide Y, leu-enkephalin and/or neurokinin A have been observed in CART-positive enteric neurons, but the degree of co-localization of CART with these substances has been low [21,38–40]. It should be underlined that the types of neuronal active substances occurring in CART-positive enteric neurons clearly depend on the animal species studied, the digestive tract fragment, and the types of enteric plexuses [21].
Due to the fact that (as mentioned above) substances co-localizing in the same neurons often play similar functions, it seems justified to list the main roles of neuronal factors observed in CART-LI esophagal enteric neurons. One of the active substances, which has been noted in CART-positive neurons during the present study was VAChT–the marker of cholinergic nervous structures. It is commonly known that acetylcholine is the main neuromediator within the ENS, which first of all is involved in the stimulation of intestinal motility and secretion [15]. In turn, VIP and nitric oxide are known as important inhibitors of intestinal smooth muscles contractions as well as gastric and intestinal secretory activity [26]. Moreover, these substances are also involved in the regulation of the blood flow in the wall of the digestive tract and immunological processes [15]. Contrary to acetylcholine, VIP and nitric oxide, functions of GAL within the ENS are not fully explained and clearly depend both on the segment of intestine, as well as animal species studied [41]. Nevertheless, it is known that GAL may regulate intestinal motility, as well as secretion of enzymes, hormones, and other neuronal factors [42,43]. The high degree of colocalization of CART with the above-mentioned substances observed in the present study strongly suggests that this peptide may play similar roles. Interestingly, contrary to previous studies [39], the present experiment has not shown meaningful colocalization of CART with substances considered as sensory factors, such as SP, CGRP, or LENK. This fact may denote that CART in the esophageal ENS, contrary to other parts of the nervous system, does not participate in the conduction of sensory and pain stimuli.
5 Conclusions
To sum up, the results obtained during the present study indicate that CART is widely distributed in enteric neurons within the porcine esophagus. The percentage of CART-positive neurons fluctuated from 45.2 ± 0.9% of all neuronal cells in the submucous plexus of the thoracic esophagus to 58.1 ± 5.0% in the myenteric plexus of the same part of this organ. Such numerous population of CART-LI neuronal cells may suggest the important functions of CART with this part of the digestive tract. In turn, the presence of CART in neurons within both myenteric and submucous plexuses indicates diversified roles of this peptide, which may apply to motility and secretory activity of this fragment of the digestive tract. Moreover, the obtained results show that CART in esophageal enteric neurons may colocalize with a wide range of other neuronal factors. The degree of colocalization of CART with a particular substance clearly depended on the fragment of the esophagus. Generally, the highest percentage of CART-positive neurons was simultaneously immunoreactive to VAChT, VIP, and/or nNOS. This fact suggests that CART is present in various types of neurons, both in stimulatory (cholinergic), as well as inhibitory (VIP– and/or NOS–positive) nervous structures, which may also indicate the multidirectional roles of CART in the physiological activity of the esophagus. Nevertheless, a full explanation of the exact functions of CART in the described fragment of the GI tract requires further studies.
Funding
This work was supported by grant No. NN401178639 of the State Committee for Science Research, Poland.
Disclosure of interest
The authors declare that they have no competing interest.