

1 Description
The term “Sussemiones” was first used in an informal way to group early and middle Pleistocene equid species with characteristic teeth [1]. Discoveries of new data, both paleontological and molecular [2,3], recommend a formal description and extend the temporal range of this group.
Sussemionus, n. subgenus.
Derivatio nominis:
In reference to the mixture of osteological characters, some of which are observed (namely) in the fossil equids from Süssenborn, Germany, and some others in extant hemiones.
Diagnosis:
Upper cheek teeth (Fig. 1) with peculiar plis caballin: multiple and/or with a very large base (Fig. 1A, Fig. 4A, Fig. 6), sometimes club-shaped (Fig. 1B). Such morphologies are unknown in extant species of Equus and in Allohippus. The enamel is often very plicated and the postprotoconal valley may be very deep. Protocones may be extremely short.

Upper premolars of Sussemionus. A. E. cf. verae, section of PIN 2998-243, Chukochya, loc. 26. B. E. hipparionoides, P4/ of Akha 100, Akhalkalaki. Illustrations of these teeth were also published in [1].
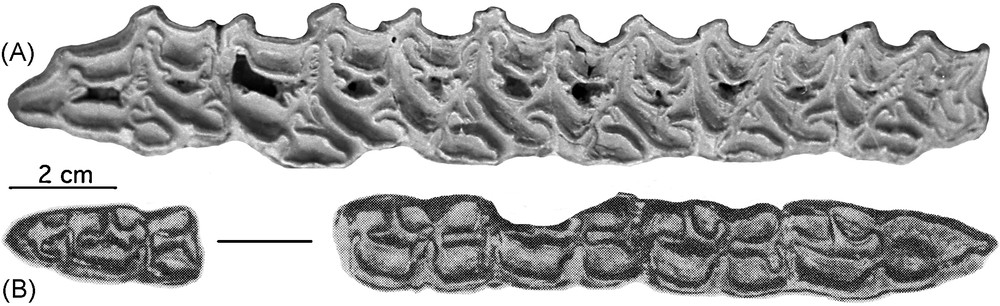
E. coliemensis. A. Left P2/-M3/ of the type skull IA 1741. B. Right P/2-M/1 and M/3 of IA 1721. Illustrations of these teeth were also published in [1].
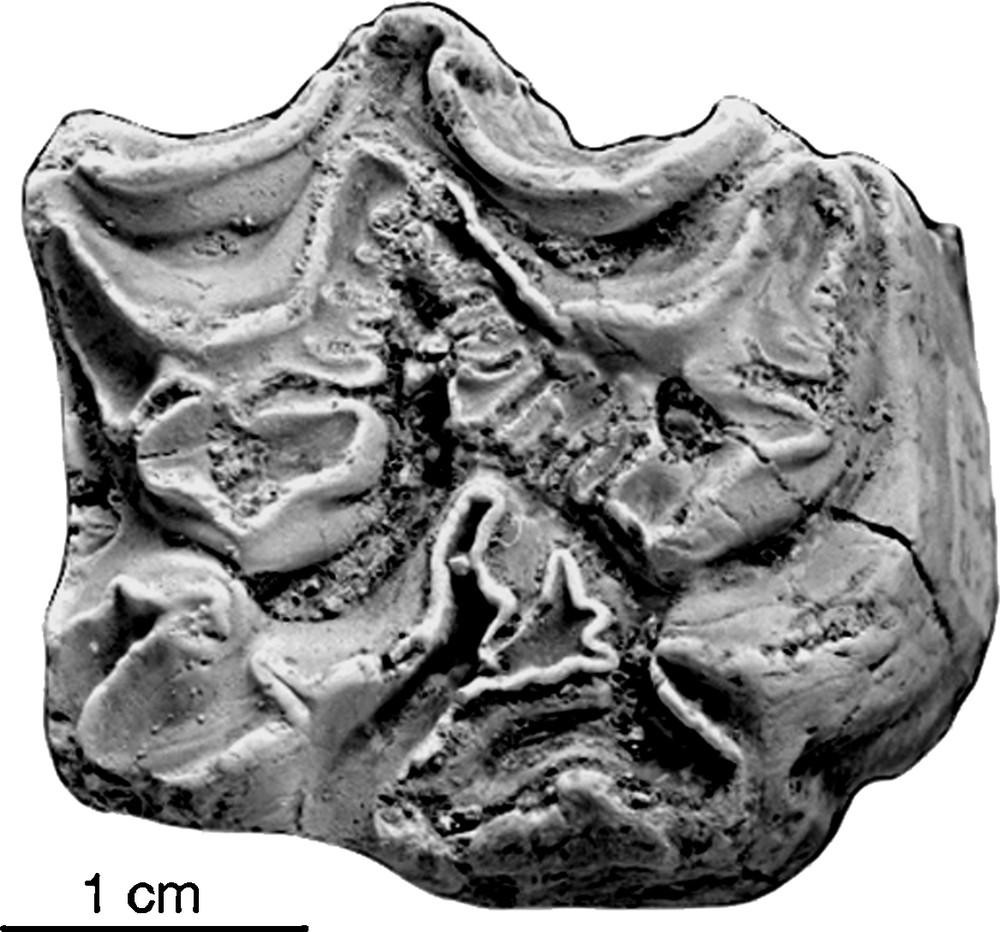
Upper premolar from the Acheulean of Gomboré II (Melka Kunturé, Ethiopia), 73-1978. Illustration also published in [1].
On the lower cheek teeth (Fig. 2), the occurence of stylids, sometimes isolated, is remarkable. Isolated ectostylids are characteristic of late African hipparions but exceptional in extant Equus. Plis protostylids on P/2 (characteristic of extant Grevy's zebras [4]) occur frequently (Fig. 2A). Plis protostylids on P/3-M/3 may be extremely developed (Fig. 2B) as well as plis hypostylids (Fig. 2C); the latter may even be isolated on M/3. The shape of the double knot of many lower premolars resembles extant hemiones, sometimes in an extreme, caricatural way: the metaconid is elongated, sometimes bilobated, the lingual valley is shallow, at times nearly absent (Fig. 2D and E). Unlike Hemiones, another particularity is the frequency of very deep vestibular valleys, on molars and even on some premolars (Fig. 2F). But the depth of the vestibular valley is very variable: associated teeth may have very deep and very shallow valleys (Fig. 2G). Both features are uncommon in extant species.
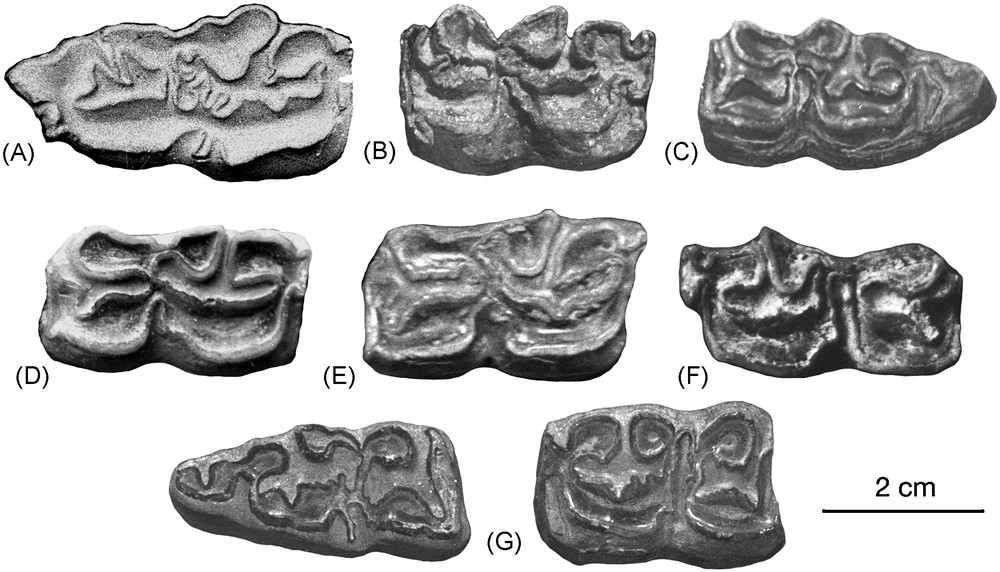
Lower cheek teeth of Sussemionus. A. P/2 of E. verae, Chukochya loc. 21, PIN 835-123. B. P/4 of E. hipparionoides, Akhalkalaki, no. 99. C. M/3 of E. cf. verae, Yukon, Old Crow loc. 9, NMC 32165. D. P/3 of E. granatensis, Venta Micena, VM 84/C3-B9-12. E. P/3 of E. verae, Krestovka, PIN 3020-47 (851-74/8). F. P/3 of E. suessenbornensis, Süssenborn, S 9281. G. Associated M/3 and M/1 of E. verae, Chukochya loc. 37, PIN 3100-333. Illustrations of B, C, D, E, and G were also published in [1]. Masquer
Lower cheek teeth of Sussemionus. A. P/2 of E. verae, Chukochya loc. 21, PIN 835-123. B. P/4 of E. hipparionoides, Akhalkalaki, no. 99. C. M/3 of E. cf. verae, Yukon, Old Crow loc. 9, NMC 32165. D. P/3 of E. Lire la suite
Type species: E. coliemensis Lazarev 1980 [5].
Origin: Kolyma, NE Siberia, Russia.
Age: late Early Pleistocene.
1.1 Description of the type material
Although not perfectly preserved, the skull IA 1741, type of E. coliemensis (Fig. 3, Table 1) belongs to Equus. The skull is as large as a Grevy's Zebra but with proportions more like Hemiones [6].
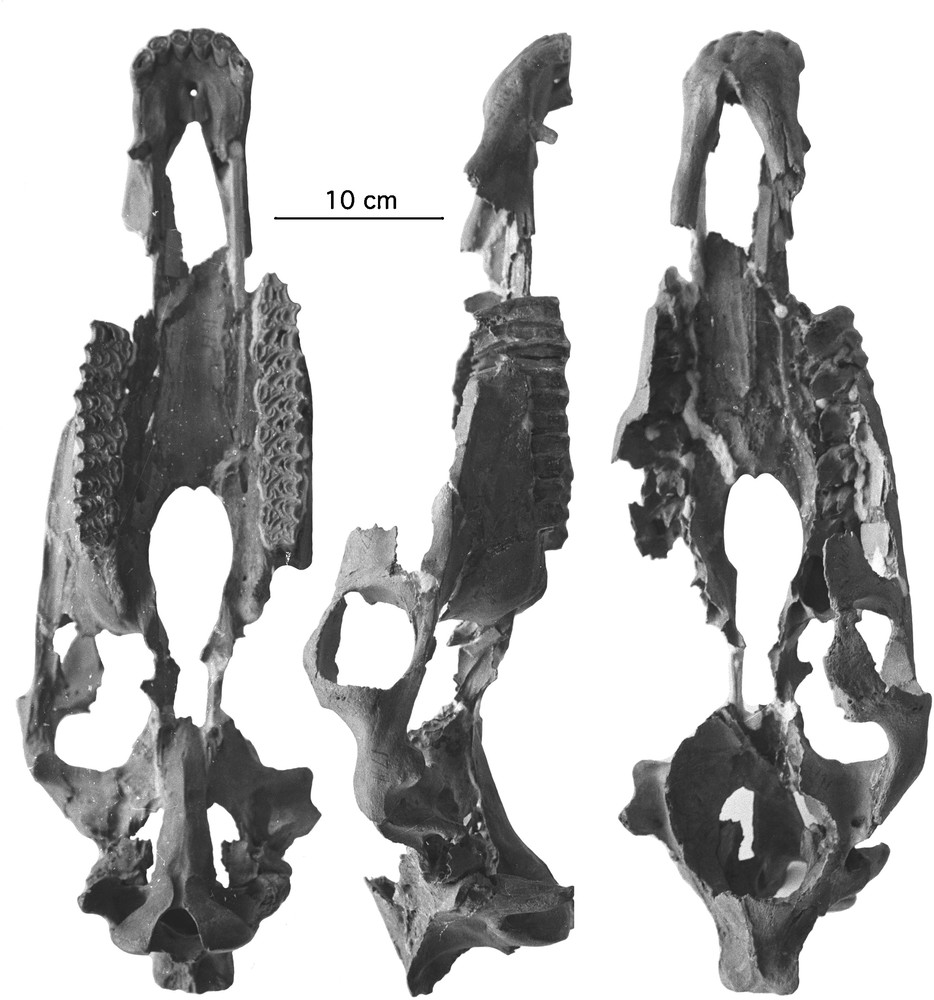
E. coliemensis skull, IA 1741. From left to right: ventral view, profile, dorsal view.
Measurements in mm of E. coliemensis skull. The numerotation of the measurements refers to the system illustrated on the web site www.vera-eisenmann.com. Approximate dimensions between brackets.
| Chukochya | ||
| E. coliemensis | ||
| IA 1741 | ||
| 1 | Basilar length | 538 |
| 2 | Overall palatal length | 282 |
| 2–5 | Palatal length s.s. | 140 |
| 3 | Vomerine length | [133] |
| 4 | Post-vomerine length | [123] |
| 5 | Muzzle length | [140] |
| 6 | Diastema | 112 |
| 7 | P2/-P4 length | [101] |
| 7bis | M1/-M2/ length | 82 |
| 8 | P2/-M3/ length | [180] |
| 10 | Greatest choanal breadth | 53 |
| 10bis | Least choanal breadth | 42 |
| 11 | Facial breadth | 164 |
| 12 | Length from Basion to anterior borders of P2/ | [398] |
| 13 | Frontal breadth | 230 |
| 14 | Bizygomatic breadth | 210 |
| 15 | Cranial breadth | 105 |
| 16 | Breadth of supra-occipital crest | 53 |
| 17 | Muzzle breadth at posterior borders of I3/ | 70 |
| 17bis | Least muzzle breadth (between the crests) | 55 |
| 18 | Greatest length | 590 |
| 19 | Height of the infra-ocular bar | 11 |
| 20 | Height of the external auditive meatus | 16.5 |
| 21 | Antero-posterior orbital diameter | 62 |
| 22 | Dorso-ventral orbital diameter | 58.5 |
| 23 | Anterior ocular line | 440 |
| 24 | Posterior ocular line | 225 |
| 29 | Breadth at the occipital condyles | 90 |
| 30 | Breadth of foramen magnum | 34 |
The upper cheek teeth of the type are plicated and have plis caballins wide at their base (Fig. 4A, Table 2). There are no associated lower cheek teeth but some were referred to E. coliemensis by Lazarev (Fig. 4B, Table 3). On the lower cheek teeth, the enamel is plicated and the hypostylid very developed on M/3.
Measurements in mm of some upper cheek teeth dentitions of various Sussemionus species. L: occlusal length. Prot L: occlusal length of the protocone. W: occlusal width. Ht: height of the crown. Additional data may be found on www.vera-eisenmann.com.
| P2/ | P3/ | P4/ | ||||||||||||
| L | Prot L | W | L | Prot L | W | Ht | L | Prot L | W | Ht | ||||
| E. süssenbornensis | Süssenborn | Type Wüst | 44.2 | 11.4 | 30 | 33.6 | 12.6 | 30 | 35.4 | 15 | 33 | |||
| E. süssenbornensis | Süssenborn | S 514 | 8.5 | 29 | 31 | 13.5 | 33 | 31.5 | 15 | 33 | ||||
| E. cf. süssenbornensis | Akhalkalaki | Akha 1285 | 36 | 14 | 32 | 27 | 30.4 | 12 | 32 | 26 | ||||
| E. coliemensis | Kolyma | IA 1741 | 39.7 | 8.3 | 27.2 | 32 | 12.7 | 30.5 | 29.6 | 12.2 | 29.6 | |||
| E. granatensis | Venta Micena | C3-87 | 38.5 | 8 | 27 | 29 | 9 | 27 | 28 | 11 | 28 | |||
| E. hipparionoides | Akhalkalaki | Akha 100 | 29 | [7.5] | 28 | 39 | 25 | 6.0 | 26 | |||||
| M1/ | M2/ | M3/ | ||||||||||||
| L | Prot L | W | Ht | L | Prot L | W | Ht | L | Prot L | W | Ht | |||
| E. süssenbornensis | Süssenborn | Type Wüst | 30.4 | 12.5 | 31.7 | 31 | 13.5 | 28.7 | 30 | 14.2 | 25.2 | |||
| E. süssenbornensis | Süssenborn | S 514 | 28 | 14 | 30 | 29 | 13.1 | 30 | 33.9 | 16.6 | 27.2 | |||
| E. cf. süssenbornensis | Akhalkalaki | Akha 1285 | 27.3 | 11.7 | 30 | 27 | 29.5 | 14.5 | 30.5 | 32 | 34 | 15 | 26 | 27 |
| E. coliemensis | Kolyma | IA 1741 | 26.6 | 11.4 | 27.9 | 26.2 | 12.3 | 26.1 | 28.4 | 12.4 | 24.8 | |||
| E. granatensis | Venta Micena | C3-87 | 26 | 8.5 | 27 | 27 | 11 | 27 | 26.5 | 8 | 22.5 | |||
| E. hipparionoides | Akhalkalaki | Akha 100 | 26 | 7.0 | 24 | 42 | 26 | 10 | 23.5 | 43 |
Measurements in mm of some lower cheek teeth dentitions of various Sussemionus species. L: occlusal length. L DN: occlusal length of the double knot. L postf: occlusal length of the postflexid. W: occlusal width. Ht: height of the crown. Approximate dimensions between brackets. Additional data may be found on www.vera-eisenmann.com.
| P/2 | P/3 | |||||||||||
| L | L DN | L postf | W | Ht | L | L DN | L postf | W | Ht | |||
| E. süssenbornensis | Süssenborn | S 9281 | 39 | 17.7 | 17 | 18 | 38 | 35 | 23 | 13 | 19.2 | 46 |
| E. cf. süssenbornensis | Akhalkalaki | Akha 1 | 42 | 16.7 | 19 | 16 | 44 | 35 | 20 | 17.5 | 18 | 55 |
| E. verae | Chukochya, loc. 21 | PIN 835-123 | 41 | 20.5 | 18 | 18.7 | [65] | 34 | 20.5 | 17.5 | 22 | 70 |
| E. granatensis | Venta Micena | VM 84 C3.B9.12 | 36.6 | 14.2 | 17.4 | 16 | 32.7 | 19.2 | 16.8 | 17.2 | ||
| E. hipparionoides | Akhalkalaki | Akha 99 | 37 | 17 | 23.5 | 33 | 18.3 | 14 | 17 | 21 | ||
| P/4 | M/1 | |||||||||||
| L | L DN | L postf | W | Ht | L | L DN | L postf | W | Ht | |||
| E. süssenbornensis | Süssenborn | S 9281 | 33 | 21 | 11.1 | 20 | 51 | 30 | 17.5 | 11 | 16 | 44 |
| E. cf. süssenbornensis | Akhalkalaki | Akha 1 | 33 | 18.5 | 15.3 | 18 | [67] | 30 | 17 | 11 | 17 | |
| E. verae | Chukochya, loc. 21 | PIN 835-123 | 32.5 | 20 | 16 | 22 | 77 | 30 | 18.3 | 16 | 19 | 67 |
| E. granatensis | Venta Micena | VM 84 C3.B9.12 | 29.5 | 18 | 13.2 | 16.8 | 28.4 | 16.8 | 10.2 | 14.5 | ||
| E. hipparionoides | Akhalkalaki | Akha 99 | 30.5 | 16.1 | 12.8 | 16.2 | 28 | 28.3 | 16.1 | 8.7 | 16.8 | 20 |
| M/2 | M/3 | |||||||||||
| L | L DN | L postf | W | Ht | L | L DN | L postf | W | Ht | |||
| E. süssenbornensis | Süssenborn | S 9281 | 30 | 17.2 | 11 | 16 | 48 | 35 | 14.5 | 16.2 | 50 | |
| E. cf. süssenbornensis | Akhalkalaki | Akha 1 | 31.7 | 16.5 | 11 | 16 | 50 | 37 | 16 | 15 | 45 | |
| E. verae | Chukochya, loc. 21 | PIN 835-123 | 29.5 | 17 | 14 | 20 | 67 | 35 | 15.5 | 14 | 17 | 69 |
| E. granatensis | Venta Micena | VM 84 C3.B9.12 | 28 | 15.6 | 8.9 | 14.3 | 33 | 15.2 | 9.3 | 13.7 | ||
| E. hipparionoides | Akhalkalaki | Akha 99 | 29 | 14.2 | 9 | 14.7 | 25 |
Other species already referred to Sussemiones – now the new subgenus (see description and illustrations in [1]):
- – E. suessenbornensis (Süssenborn, Germany), E. cf. suessenbornensis (Akhalkalaki, Georgia; Cueva Victoria, Spain), E. verae (NE Siberia), E. cf. verae (Alaska, Yukon). The metapodials are large and robust;
- – E. granatensis (Venta Micena, Spain), E. hipparionoides of Akhalkalaki, the very poorly defined E. altidens and the slightly better founded E. marxi of Süssenborn. Teeth and/or metapodials of this group are found in the British Forest Beds (Trimingham), and in Spain (Cueva Victoria, Cullar de Baza, Huescar). The metapodials are slender.
1.2 New data
Orlando et al. [2] analysed some late Pleistocene samples from Proskuriakova cave (Khakassia, southwest Siberia, Russia) believed to belong to a kind of E. hydruntinus. Unexpectedly, instead of an E. hydruntinus or some hemione, they found a new clade of Equus with no extant relative, and distinct from E. hydruntinus.
A more detailed osteological study of the fossils from Proskuriakova cave confirmed the biomolecular findings: the Equus from Proskuriakova is neither an hydruntine nor an hemione; it shows dental features of Sussemiones [3] (description in press). In the course of the same study, it was discovered that another late Pleistocene Equus, believed to belong to a hydruntine, should also be referred to Sussemiones. It was excavated at the Mousterian site of Tsopi, Georgia.
2 Discussion and conclusions
2.1 Origin of Sussemionus
In the late Blancan of Arizona, there is a very small species [7] reminding of Sussemionus (Fig. 5A). Another, much larger species, with marked Sussemionus characters (Fig. 5B) was found in Alaska inside probably Pliocene deposits (more than 2 Ma, A. Sher pers. comm.). It seems thus that, like most equids, Sussemiones originated in North America.
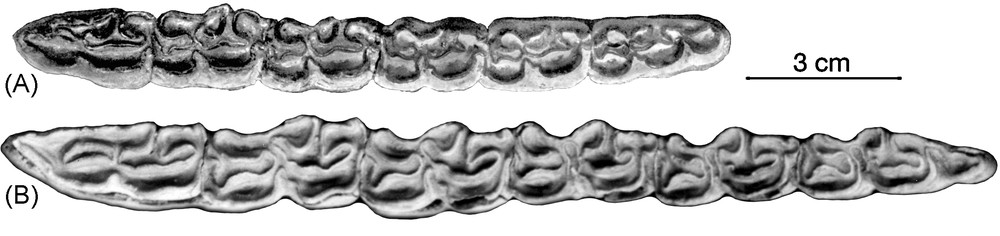
Lower cheek teeth series tentatively referred to Sussemionus. A. AMNH 116502, Dry Mountains, Arizona, adapted from Azzaroli and Voorhies 1993. B. Lost Chicken, Alaska. Illustrations of these teeth were also published in [1].
2.2 Geographical and chronological ranges
The subgenus Sussemionus was very successful, judging by its geographical distribution from Arizona to possibly Ethiopia (Fig. 6), and its chronological range from ca 2 Ma to about 45–50 ka (Proskuriakova cave).
2.3 Environments
Dental and skeletal morphologies of extant Equus allow one to make assumptions as to the environment of fossil equids. Usually, a simple enamel is observed in species feeding on hard vegetation, while a more complicated enamel is found in species feeding on a softer one [8, p. 98]. Slenderness and cursorial proportions are linked to open landscapes, while robustness and ‘graviportal’ proportions are observed in opposite cases [9].
The extremely rich collection of equids from Venta Micena (Spain) include no other species than E. granatensis. This equid had a simple enamel, slender limb bones, and cursorial proportions [10]. On the other hand, in Chukochya, from where E. coliemensis and E. verae (possibly a junior synonym) were described, there is no evidence of a slender species. E. coliemensis had a very plicated enamel and robust limb bones.
Things are more complicated for the other findings where both slender and robust equids were excavated: the time encompassed by Süssenborn deposits is very long; Cueva Victoria may have been equally heterogeneous. It seems that Akhalkalaki is the only place where a robust form (E. cf. suessenbornensis) and a slender one (E. hipparionoides) have coexisted, at least in a broad paleontological sense.