

1 Introduction
The hammerhead sharks are grouped in the monophyletic family Sphyrnidae, comprising eight nominal extant species in two genera (i.e. Sphyrna and Eusphyra) as follows [1]: Sphyrna lewini, S. mokarran, S. tiburo, S. tudes, S. zygaena, S. media, S. corona and Eusphyra blochii.
This family is known since the Lower Miocene of Portugal [2], and is closely related to the family Carcharhinidae based on morphological [3] and molecular features [4–6].
Some groups of sharks can be identified by tooth characters related to the shape of the crown and the root, number of rows and arrangement of nutritive foramina [7,8].
These characters are especially important to identify fossil records, sometimes species level [9]. This importance is justified by the difficulty to find other fossilized body parts in sharks that are not teeth and dermal denticles. Furthermore, the dental characters in embryos remain unknown, as well as intra- and inter-specific comparisons within the family Sphyrnidae.
The study of fossil forms allows us to reconstruct past diversity patterns [9], and, compared to living forms, collaborates to understand the phylogenetic position and evolutionary traits of many groups of sharks.
The teeth in sphyrnids are often confused with carcharhinids teeth, especially considering the identification of fossil records. It is known that sphyrnids teeth are very similar to Rhizoprionodon, Loxodon and Scoliodon [10].
The ontogenetic heterodonty is weak among sphyrnids [3], and the sexual heterodonty is hardly developed and there are brief descriptions of the teeth in the species of Sphyrnidae [3,11]. However, the teeth in extant species are usually illustrated in only one arbitrary view and examining just a few specimens.
To many authors, the teeth in sharks are divided according to their position, as follows: symphisials, parasymphyseals, anteriors, intermediates, laterals and posteriors [12–15]. For the Carcharhiniformes, the tooth nomenclature has some subdivisions [3] as follows: mesials, symphisials, anteriors, anteroposteriors, anterolaterals, lateroposteriors, laterals and posteriors.
However, this classification is rather subjective than useful, especially to establish tooth homologies. This lack of objectiveness is due to the tooth type terminology, which is based mainly on (a) the general area of the jaws and (b) relative terms to define each tooth type [12,15].
Therefore, the aims of this study are:
- (a) to analyze intra-specific dental differences between early embryos of the three studied species of Sphyrnidae;
- (b) to study dental characters that could be lost in later ontogenetic stages;
- (c) to describe new characters in extant sphyrnid teeth in different anatomical views;
- (d) to suggest a more functional and objective classification of tooth types based on morphological patterns in S. tudes, S. tiburo and E. blochii.
2 Material and methods
Specimens of S. tiburo, S. tudes and E. blochii were examined in early embryonic stage, including males and females, forming the focus of this study. Some juveniles of the same species were also examined just to verify the data in the literature and possible changes in tooth morphology. Early embryos are considered here as specimens with the umbilical cord or, at least with the umbilical hole. All the embryos (males and females) examined have a minimum of 20 cm of total length as they have tooth developed enough to be observed.
The teeth of some specimens were collected and prepared with dermestid beetles to separate them from the organic material. The teeth were cleaned in a solution with 10% of sodium hypochlorite for about 2 h and washed with water to clean the rest of the eventual organic material. A few teeth of every row in one half of each jaw were collected, in order to preserve the rest of the rows. Each tooth type were photographed and schemed in lingual and labial views (and in some cases in additional views, such as lateral, basal or apical) to facilitate the observation of morphological features.
The following dental characters were studied: central cusp (cc), central foramen (cf), crown-root junction (crj), distal blade (db), distal cusplet (dcl), distal heel (dh), mesial cusplet (mcl), mesial blade (mbl), mesial heel (mh), notch (no), root (ro), root foramina (rf) and transversal groove (tg). The morphological nomenclature follows Cappetta (1987), and is complemented by Compagno [3].
The material used is deposited in the National Museum of Natural History/Smithsonian Institution (NMNH), California Academy of Sciences (CAS), Musée National d’Histoire Naturelle in Paris (MNHN), Universidade do Estado do Rio de Janeiro (UERJ) e Museu Nacional da Universidade Federal do Rio de Janeiro (MNRJ).
The number of specimens examined is indicated in parenthesis, preceded by its number for each species as follows:
- S. tiburo (14 specimens): MNRJ 9968 (1), 37342 (2), 37343 (1), 37344 (1), 37345 (3), 37346 (2), 37347 (3); CAS 11224 (1); NMNH 147793 (1), 116888 (11), 155964 (17 embryos), 222050 (1), 395407 (1);
- S. tudes: MNRJ 44660 (8); NMNH 87682 (1), 120751 (1); CAS 52875 (1);
- E. blochii: NMNH 195846 (2), 197512, 205342 (5), 222045 (1), CAS 39180 (1), 58058 (1); SU 13815 (1), 14495 (2), 30899 (1), 41989 (2).
3 Results
No sexual or ontogenetical heterodonties were found in the embryos of the studied species. However, when compared to juvenile data in the literature [11], some differences were observed.
All teeth in embryos are of cutting-type (sensu Cappetta, 1987), which also occurs in the related taxa of Carcharhinidae.
The tooth types observed in each sphyrnid species are asymmetrical (Fig. 1) or symmetrical (Fig. 2).
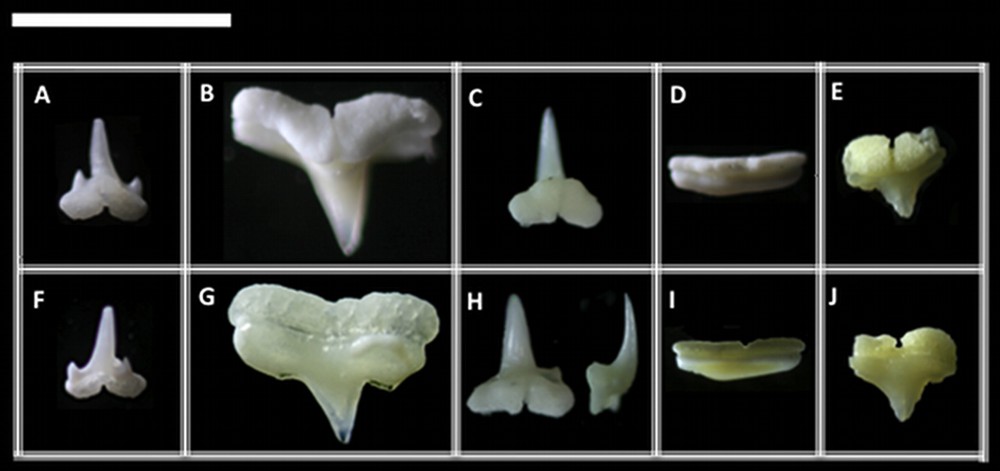
Symmetric groups of teeth: (A) with cusplets, (B) without cusplets, (C) inclined distally with a prolonged crown, (D) with a vestigial cusp and (E) with a developed root. Photographs were taken in lingual (A to E) and labial (F to J) views for each respective tooth. Scale bar = 5 mm.

Asymmetric groups of teeth: (A) inclined with cusplets, (B) inclined and well cuspidated, (C) inclined and vestigial cuspidated. Photographs were taken in lingual (A to C) and labial (D to F) views for each respective tooth. Scale bar = 5 mm.
The tooth types are described here for the first time as follows:
3.1 Sphyrna tiburo
Type 1 (Figs. 1A, F and 3): the mesial and distal blades of the central cusp are triangular in shape and labio-lingually curved. The distal and mesial heels are differentiated to form, respectively, the distal and mesial cusplets, which are pointed. In adults, including fossil records, these cusplets loose their sharp tips, which become blade-like in shape. This modification suggests that the mesial and distal heels are homologous, respectively, to the mesial and distal cusplets. Such heels are also named as shoulders [13]. The cusplets are separated from the central cusp by a deep notch.
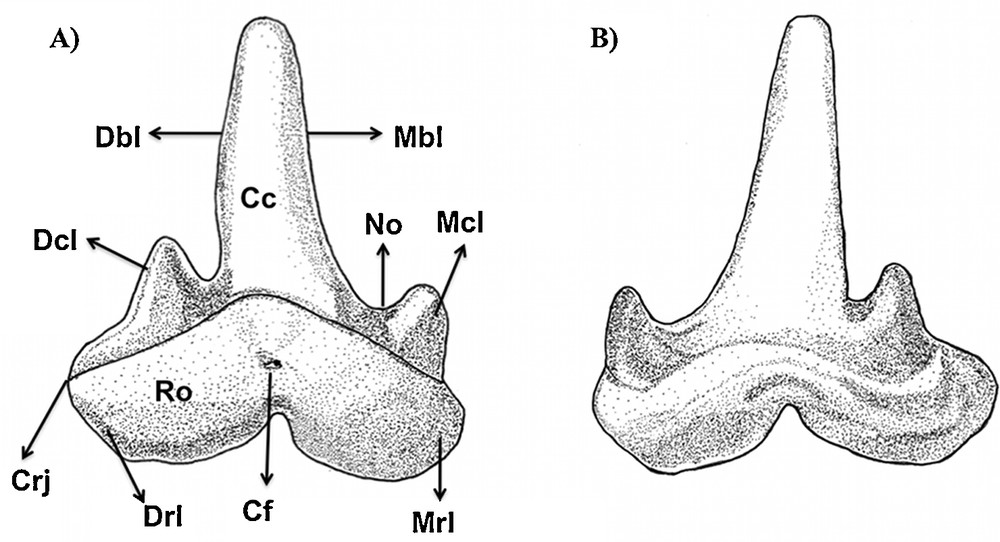
Type 1 tooth of Sphyrna tiburo in (A) lingual and (B) labial views.
The root foramina are not visible in labial view as in other teeth (showed forward), as they are covered by the elevation of the basal ledge. However, the root foramina can be seen in the basal and lingual views of the root.
The mesial and distal attachments of the root are developed. The crown/root junction is concave in labial view. Below this junction and in labial view, there is a developed depression. In lingual view, the root presents a developed center-lingual protuberance. The transversal groove is well visible in lingual view, which passes labio-lingually through the basal surface of the root, ending in the central foramina.
This tooth type is relative to the mesial or anterior teeth, being as small as the posterior ones (sensu Compagno, 1988) of sphyrnids.
Type 2 (Figs. 1B, G and 4): the tooth has a slightly sigmoid mesial blade, and its central cusp is inclined. The distal blade is smooth and separated from the cusp by a well-defined notch. In labial view, the region of the notch presents a subtle depression.
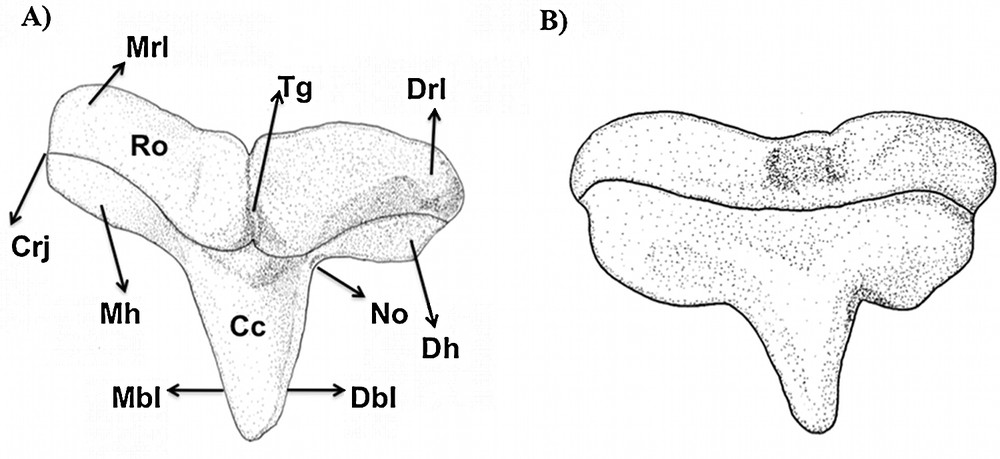
Type 2 tooth of Sphyrna tiburo in (A) lingual and (B) labial views.
In labial view, the root presents many foramina organized transversally. The mesial and distal attachments of the root are more developed and convex than in tooth type 1. These lobes extend to the middle portion of the tooth in lingual view, to form a protuberant root in lingual view. The transversal groove is present and well developed in this same view, where the central foramen is inserted. In labial view, the root is more flattened and a series of foramina can be observed.
This tooth type is relative to the anterior to lateral teeth (sensu Compagno, 1988).
Type 3 (Figs. 2C, F and 5): the cusp is very reduced and inclined. Its height is short, reaching less than the height of the crown. There is a deep notch separating the cusp from the distal heel, which presents a characteristic labio-lingual depression to form the blade-like shape. The mesial heel is lost in this tooth type in comparison to the others. The crown/root junction is slightly straight, and the root is flattened without apparent foramina. The transversal groove and the central foramina are subtle in lingual view.
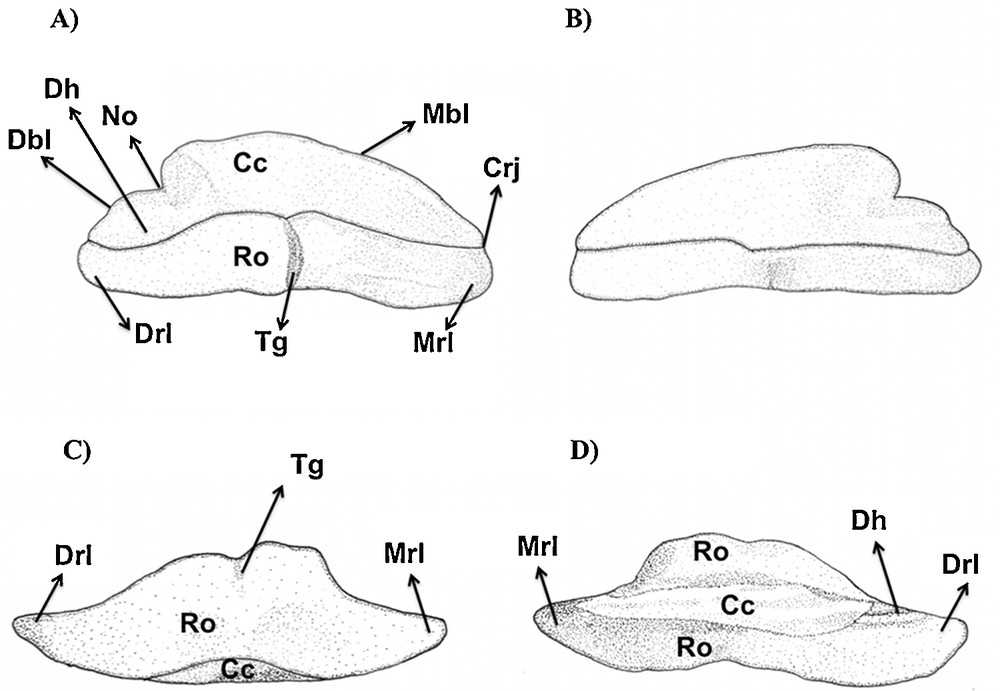
Type 3 tooth of Sphyrna tiburo in (A) lingual, (B) labial, (C) basal and (D) apical views. Type 3: this tooth is similar to the type 3 of S. tiburo. Type 4: no tooth without a central cusp (like in S. tiburo) was detected in this species. Nevertheless, it should not be discarded in big size specimens by sexual or feeding consequences.
This tooth type is morphologically intermediate to patterns 2 and 4 in S. tiburo. The particular differences in relation to the latter types are related to the very short cusp with a small distal tip and the absence of a mesial heel. This tooth type is relative to the lateral to posterior teeth (sensu Compagno, 1988).
Type 4 (Figs. 1D, I and 6): the central cusp and heels are absent, with a continuous blade, which is more or less convex.
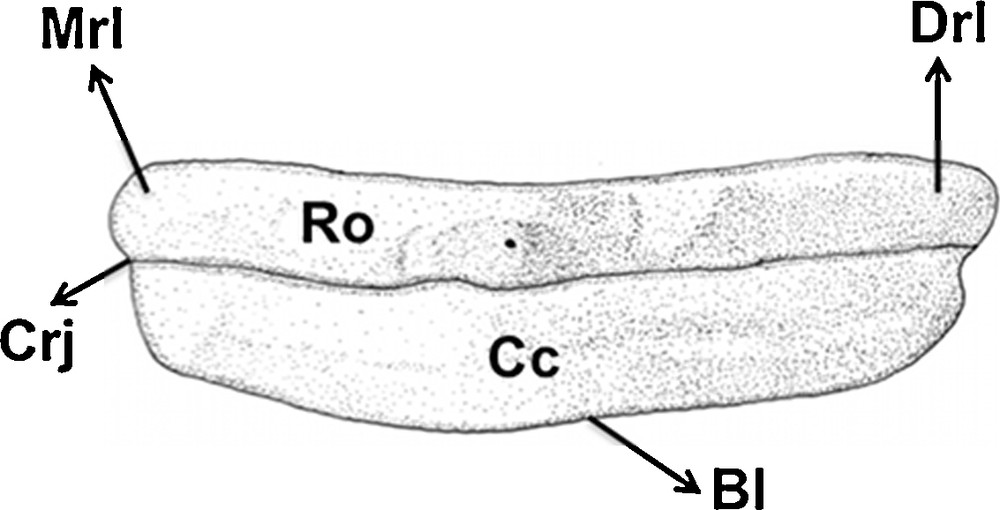
Type 4 tooth of Sphyrna tiburo in lingual view (equal to the labial view).
The crown height is short, with about two times the root height. The base of the root is straight, with one central foramen in the middle portion in lingual view. The transversal groove is obscure. Both lingual and labial views are very similar. This tooth type is relative to the posterior teeth (sensu Compagno, 1988).
3.2 Sphyrna tudes
Type 1 (Figs. 2A, D and 7): the mesial and distal blades of the central cusp are slightly sigmoid and turned towards the corner of mouth.
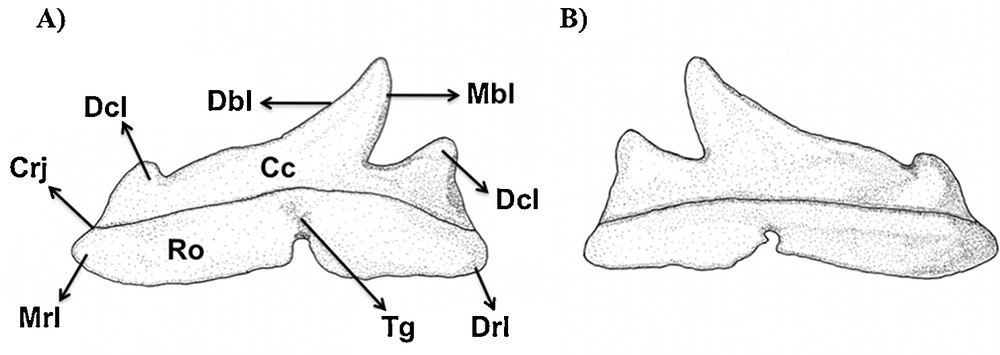
Type 1 tooth of Sphyrna tiburo in (A) lingual and (B) labial views.
A distal heel is present. However, the mesial heel is differentiated to form a mesial cusplet, which is pointed. In adults, the cusplet looses its sharp tip, which changes to a rounded blade-like shape. This modification suggests that the mesial heel and the mesial cusplet are homologous. The cusplet is separated from the central cusp by a deep notch.
The root foramina were not observed in any view.
The root is relatively short and its mesial and distal edges are less developed when compared to S. tiburo and E. blochii. The crown/root junction is concave in labial view. In lingual view, the root presents a developed center-lingual protuberance. The transversal groove is well visible in lingual view, presenting the central foramen.
This tooth type is relative to the mesial to anterior teeth (sensu Compagno, 1988).
Type 2 (Figs. 1C, H and 8): this type showed to be characteristic for the lower jaw of this species. It is very similar to the type 1 of S. tiburo, except by the much taller central cusp, which can be about one half times taller than in S. tiburo. Additionally, the cusp is strongly labio-lingually inclined, with a deep transversal groove throughout the root base. It can eventually present a smooth cusplet-like mesial heel in the most anterior rows. This tooth type is relative to the mesial to anterior teeth (sensu Compagno, 1988).
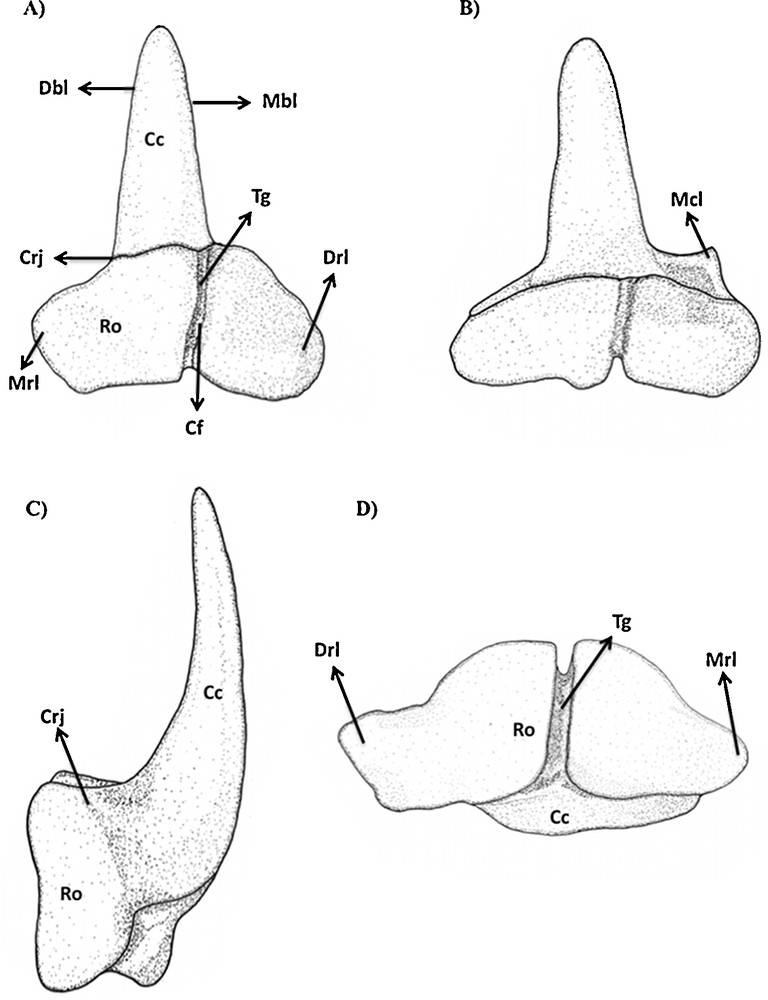
Type 2 tooth of Sphyrna tudes in (A) lingual, (B) labial, (C) lateral and (D) basal views.
3.3 Eusphyra blochii
Type 1 (Figs. 1E, J and 9): the mesial and distal blades of the central cusp are straight and triangular in shape. The distal and mesial heels are absent. The mesial and distal attachments are the most developed and convex in lingual view compared to any other tooth in the other examined species. They extend to the middle portion of the tooth in lingual view to form a deep transversal groove, which presents a central foramen in its middle portion.
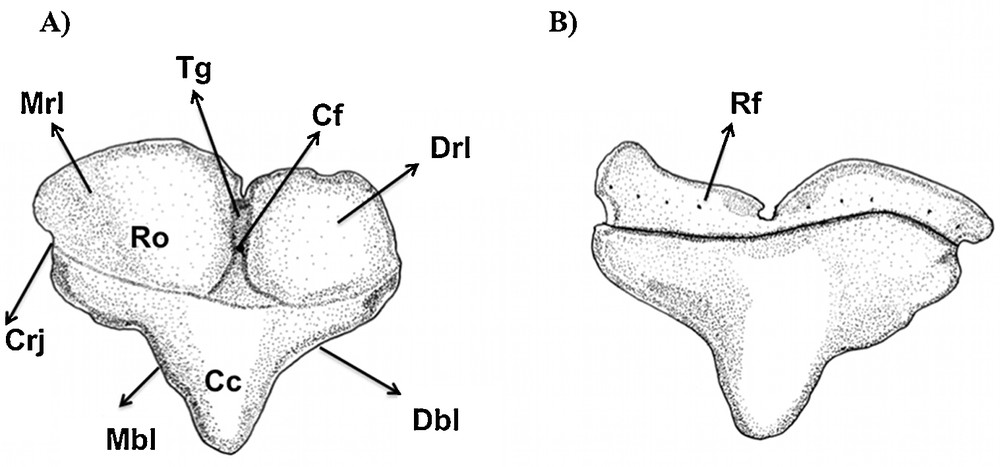
Type 1 tooth of Eusphyra blochii in (A) lingual and (B) labial views.
In labial view, the root is smoothly flattened, presenting a row of foramina. The elevation of the basal ledge covers the root foramina in lingual view, but they also are observable in basal and labial views of the root.
The crown/root junction is curved in lingual and labial views.
This tooth type is relative to the mesial teeth (sensu Compagno, 1988).
Type 2 (Figs. 2B, E and 10): the tooth has a slightly sigmoid mesial blade, and its central cusp is inclined. The distal blade is separated from the cusp by a well-defined notch. In labial view, the region of the notch presents a subtle depression. The mesial heel is absent.
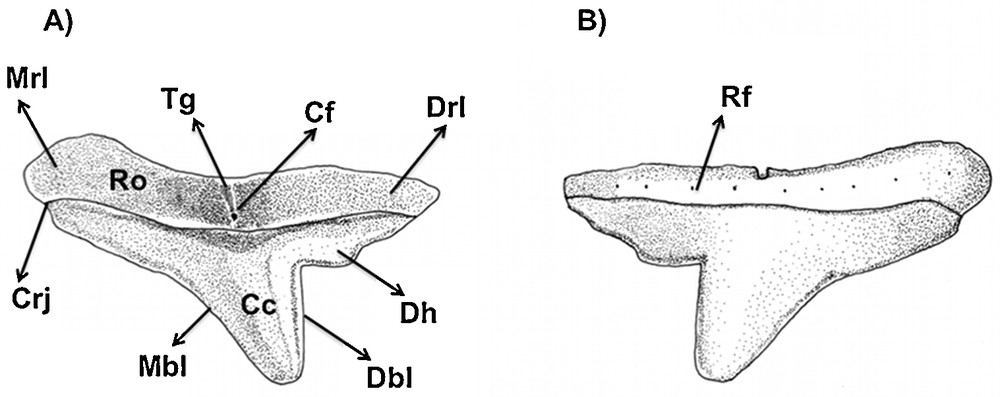
Type 2 tooth of Eusphyra blochii in (A) lingual and (B) labial views. Types 3 and 4: similar to S. tiburo.
In labial view, the root presents a row of foramina organized transversally. The mesial and distal attachments of the root are developed and convex as in the same tooth type in the other observed species. The transversal groove is present and well clear in this same view, where the central foramen is inserted. In lingual view, the root is the most prominent compared to other tooth types in the studied species. In labial view, the root is more flattened and a series of foramina can be observed.
This tooth type is relative to the anterior to the lateral teeth (sensu Compagno, 1988), and is very similar to the one observed in S. tiburo. Nevertheless, it presents a lesser prominent root.
4 Discussion
Frequently, only one tooth view is considered to describe teeth in species of Sphyrnidae, or even in its most related family Carcharhinidae. Although the tooth descriptions, including fossil sphyrnids [15–17] and of many other genera [18,19], there is a scarcity of publications for living taxa, which frequently show the teeth in labial or lingual view. This lack of important data has been supporting the neglect of teeth as a tool of comparison among living and fossil species. Furthermore, embryos of sphyrnids presented tooth characters that are modified in other developmental stages, as showed in the present study, especially in the anterior teeth.
The tooth positions often depend on the identification of the adjacent ones, which makes their distinction ambiguous in many cases [20]. When just few teeth are available, this recognition is even harder, which often happens in fossils consisting of isolated teeth.
The use of tooth types as suggested in this paper could contribute to define what kind of particular teeth are found in every species and its transformations from the early embryo to advanced stages of development. The study of early embryos allowed observing characters that are modified in juveniles or adults. The results showing tricuspidated teeth in early embryos of sphyrnids corroborate such hypothesis. There are a few works [3,11] presenting the upper and lower tooth sets of sphyrnids, and even for related genera [10]. Therefore, the description of new early tooth characters and tooth types were rather the aims of this study than the reconstruction of the tooth sets.
Therefore, the reconstruction of such tooth sets were not the aim of the present work, but the description of new tooth characters and early tooth types.
The presence of cusplets (observed in S. tudes and S. tiburo) is known in the genera Scoliodon, Rhizoprionodon, Negaprion and Triaenodon, and the secondary loss of such cusplets is known in the first two genera [3,10]. These genera are phylogenetically related with the monophyletic family Sphyrnidae [3]. The presence of root foramina in the studied species is a character shared with many fossil carcharhinids [13].
To minimize possible confusions with fossil or extant carcharhinids and sphyrnids, labial and lingual views must be considered, in addition to the examination of dental series instead of just isolated teeth. The identification of sphyrnids based on isolated teeth is not encouraged for specific level. Nevertheless, the presented characters in the anterior teeth give a new perspective to understand what carcharhinid-like characters are maintained in sphyrnids.
5 Conclusions
The present study shows basically five groups of symmetric teeth and three groups of asymmetric teeth, examining males and females in early embryonic stage of S. tudes, S. tiburo and E. blochii, comparing with the known data about juveniles and adults. The asymmetric groups (and positions sensu Compagno 1988) are: inclined with cusplets (S. tudes) in mesial position, inclined and well cuspidate (S. tiburo, S. tudes and E. blochii) in anterior to lateral positions, inclined vestigially cuspidated in posterior position (S. tiburo, S. tudes and E. blochii).
These groups show that the differences in the tooth morphology of S. tiburo, S. tudes and E. blochii are basically related to the most anterior teeth in both jaws (i.e. type 1 and 2). Lateral and posterior teeth (sensu Compagno 1988) present the same basic morphology.
The root foramina, cusplets and shape of the central cusp and root presented to be good sources of characters to distinguish the studied sphyrnids by sets of teeth.
While the most posterior teeth presented a clear similarity in the studied features [3,11], distinguishable characters were observed in the most anterior teeth, suggesting that such characters could be secondarily lost in juveniles and adults. Moreover, these features may help comparisons with fossil forms in the family Sphyrnidae, as well as to understand the changes occurring during their development. This is the first step to study characters that may collaborate to clarify differences among fossil tooth records of the closely related sphyrnids and carcharhinids.
Disclosure of interest
The authors have not supplied their declaration of conflict of interest.
Acknowledgements
We thank Dr Tomio Iwamoto (CAS), Dr Lynne Parenti (NMNH) and Jeff Williams for providing working space and the necessary facilities in the respective fish collections, and the Coordenação de Pessoal de Nível Superior (CAPES) for providing funds for this research. In addition, we are grateful to the reviewers for their valuable critics to this work.