

Abridged English version
We have built a process-based model of plant succession for simulating the growth of a natural vegetation during a long period of fallow. Vegetation is described as a mixture of species competing for light and soil nitrogen. Shoots are distributed among horizontal layers and randomly distributed in the horizontal plane. Each species has four organs: leaves, stems, roots, and reproductive parts. The model simulates growth and biomass production of these organs for each species, and carbon and nitrogen cycles through soil and plants. It uses meteorological input data with a daily time step. Simulated processes are photosynthesis, growth and maintenance respiration, carbon allocation, nitrogen absorption, fixation and remobilisation, organ mortality, seed dispersion and germination. Some processes as photosynthesis are computed on an hourly basis, while seed dispersal occurs only once a year. This model has been applied to tropical fallows in agroecosystems with potatoes at high elevation (3200 to 4200 m) in the Venezuelan and Bolivian Andes, as part of a European project called TROPANDES.
The simulated growth of each species in isolation follows an S curve that may be fitted by a pseudo-logistic equation. The plateau of this curve results of an equilibrium between carbon input by photosynthesis and carbon losses through respiration and senescence. Vegetation dynamics and ecosystem functioning strongly depend on initial conditions such as seed bank and seed rain, and on water and nitrogen resources. In spite of this sensitivity, the model, with six species only in its present version, is able to reproduce relatively well the successional dynamics in Venezuela with pioneer species such as Rumex acetosella, secondary species such as Lupinus sp., and late species (shrubs of genera Hypericum, Baccharis and Acaena) and giant rosettes of the genus Espeletia.
A sensitivity study has revealed the predominant role of the legume species in this system that is strongly limited by nitrogen. When available soil nitrogen is progressively increased, community production increases to a maximum and then decreases at higher nitrogen availability, which seems odd at first sight. The explanation may be found when looking at the simulated growth of individual species. The increase in soil nitrogen results in a progressive suppression of the legume species, which is a fast growing species, and its progressive replacement by shrubs. Why are shrubs successful competitors to a fast-growing legume? They have a slower growth but having a higher size they are able, given time, to shade the legume and thus to outcompete it. Such species replacement may occur in other similar cases when several factors interact. It is nearly impossible to predict the behaviour of such systems by the sole intuition, which makes the use of models quite useful.
1 Introduction
La modélisation est une technique qui s'est beaucoup développée avec l'apparition des ordinateurs. Elle permet de résumer les connaissances disponibles sous forme d'équations et d'analyser le comportement de systèmes complexes difficiles à étudier autrement. Dès la fin des années 1960, le Club de Rome avait commandé une étude cherchant à prévoir l'évolution de la population humaine, en examinant les cinq facteurs de base qui déterminent, et donc limitent, sa croissance : population, production agricole, ressources naturelles, production industrielle, et pollution [1]. Cette étude avait montré que la population humaine arrivait nécessairement à une stabilisation, plus ou moins rapide selon le poids accordé aux divers facteurs, avec, dans certains cas (pollution importante), une réduction dramatique. Les auteurs insistaient sur le fait qu'il n'est pas possible de prévoir le comportement de leur modèle avec plusieurs facteurs en interaction. Ce modèle a été fortement critiqué, notamment parce qu'il traitait le monde globalement sans prendre en compte la diversité des situations géographiques, politiques et économiques. Par la suite, il a été détaillé en prenant en compte plusieurs grandes régions, mais les grandes conclusions sur la nécessité de limiter la croissance sont restées inchangées.
Notre étude a une portée beaucoup plus modeste, cherchant à prédire l'évolution de la végétation naturelle qui s'implante après abandon d'une culture (appelée succession secondaire en écologie). De nombreuses espèces apparaissent, mais dans un ordre relativement défini, avec des espèces pionnières, puis secondaires, puis de fin de succession. Les caractéristiques de ces espèces affectent évidemment les propriétés globales de la végétation, comme la production de biomasse. Nous allons montrer qu'il n'est pas possible de prédire par l'intuition comment cette production peut évoluer en fonction des conditions extérieures.
2 Matériel et méthodes
Nous allons distinguer le modèle qui a servi à cette étude et les conditions particulières auxquelles il a été appliqué ici.
2.1 Les hypothèses du modèle
Le modèle Faprom (FAllow PROduction Model) a été développé pour simuler la croissance de la végétation naturelle au cours de la jachère dans des systèmes de culture à jachère longue (10 ans en moyenne) caractéristiques des hautes Andes tropicales, et étudiés notamment au Venezuela et en Bolivie dans le cadre du projet européen TROPANDES [2]. On en trouvera une description détaillée avec toutes les équations et les paramètres dans [3].
La végétation est décrite comme un mélange d'espèces en compétition pour la lumière et l'azote. Les parties aériennes occupent plusieurs strates horizontales dans lesquelles les feuilles sont distribuées au hasard. Chaque espèce comporte quatre organes : feuilles, tiges, racines et organes reproducteurs. Le modèle prédit la croissance et la production de biomasse de chaque espèce, et prend en compte les cycles du carbone et de l'azote à travers le sol et les plantes. Il utilise un pas de temps journalier, avec comme entrées des variables météorologiques. Il simule divers processus : photosynthèse, respiration de croissance et d'entretien, allocation du carbone, absorption, fixation et remobilisation d'azote, exsudation racinaire, sénescence des tissus, et dispersion et germination des graines (Fig. 1).

Processus simulés dans le modèle Faprom.
L'action de la température n'est pas modélisée, car ses variations saisonnières sont assez faibles au voisinage de l'équateur. Les espèces sont en compétition pour la lumière et l'azote du sol, et ont un fonctionnement différent selon qu'on est en période sèche ou humide, mais sans prise en compte explicite de la compétition pour l'eau. La hauteur maximale de chaque espèce est fixée au départ et la hauteur réelle est proportionnelle à la biomasse (ce qui pourrait être amélioré). La hauteur conditionne la compétition pour la lumière : les plantes de grande taille portent ombrage aux plantes plus petites et sont donc favorisées, mais leur croissance initiale est plus faible, car elles doivent investir dans les organes de soutien (tige, tronc et branches pour les ligneux). Dans le modèle, les organes végétaux sont répartis en strates horizontales de 10 cm d'épaisseur, de façon homogène sur la hauteur.
L'absorption d'azote est prise proportionnelle à la longueur racinaire de chaque espèce et à la quantité d'azote disponible dans le sol, avec un maximum qui dépend de la croissance potentielle (c'est-à-dire non limitée en azote) et de la concentration en azote des tissus en croissance (prise constante par organe pour une même espèce). Alors que toutes les espèces simulées ont le même métabolisme photosynthétique (dit en C3), on distingue une légumineuse qui a la capacité à fixer symbiotiquement l'azote moléculaire de l'air. Cette fixation a un coût respiratoire additionnel par rapport aux espèces non fixatrices tirant leur azote du sol. Ce coût est évalué, d'après les données de la littérature [4], à 20% du carbone assimilé en même temps que l'azote fixé, ce qui revient pratiquement à doubler la respiration de croissance pour une espèce tirant tout son azote de la fixation.
La Fig. 2 montre comment se fait la circulation du carbone dans les plantes. Une fraction de la photosynthèse brute est utilisée pour la respiration de croissance, l'entrée de carbone dans la plante est donc (. Une partie de ce flux de carbone est utilisée pour la respiration d'entretien, le reste va servir à la croissance de la biomasse B. Deux autres flux tendent à diminuer la valeur de B : la sénescence S, qui alimente la nécromasse sur pied D, et la dispersion des graines ; un autre flux tend à l'augmenter, , qui représente l'apport de graines sur la parcelle considérée. Enfin, un flux F fait passer le carbone de la nécromasse sur pied D à la litière L (nécromasse au sol). La décomposition de cette litière et de la matière organique du sol fournit l'azote minéral nécessaire à la croissance. La circulation de l'azote dans la plante se fait selon un schéma analogue à celui de la Fig. 2, avec l'addition pour la légumineuse d'un terme de fixation symbiotique et la prise en compte d'une retranslocation d'azote au moment de la sénescence.
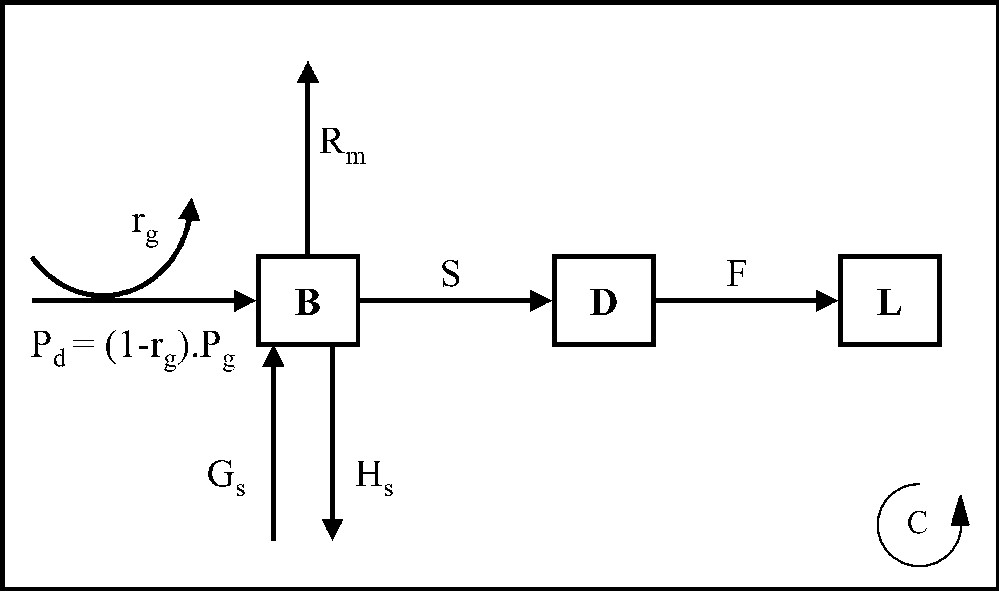
Représentation de la circulation du carbone dans les plantes.
Le modèle a d'abord été développé en Fortran puis converti en Vensim, un langage de programmation graphique de systèmes dynamiques fonctionnant en C. Il comprend 40 équations et huit paramètres pour caractériser le cycle de l'azote du sol et 48 paramètres par espèce. Sur ces 48 paramètres, 29 ont été mesurés directement sur le terrain, quatre proviennent de revues bibliographiques, neuf d'études indépendantes de ce travail ; seuls six paramètres ont été ajustés : quatre pour caractériser la mortalité (non mesurée) de l'espèce en saison sèche, et deux pour caractériser les conditions initiales (stock et pluie de graines). Il faut environ une seconde pour simuler une année de jachère sur un micro-ordinateur récent, ce qui autorise de nombreuses simulations.
2.2 Le système de culture au Venezuela
Notre modèle a une portée générale, mais il comporte de nombreux paramètres qui doivent être déterminés pour chaque application. Nous décrirons la situation dans le site vénézuélien qui est le mieux documenté [5]. Le village de Gavidia (8°35′N, 70°52′W) est situé entre 3200 et 3800 m d'altitude, dans une ancienne vallée glaciaire de la Sierra de Mérida. À la station météorologique située à 3400 m, les précipitations moyennes sont de 1300 mm an−1 (avec une saison sèche de 4 à 6 mois), la température moyenne annuelle de 8,4 °C et le rayonnement global journalier moyen est de 15,8 MJ m−2. Le système de culture comporte une jachère de 3 à 20 ans (la moyenne se situe vers 10 ans), suivie d'une culture de pomme de terre pendant un ou deux ans, et éventuellement une culture autre (blé, carotte, ail) avant la jachère suivante. Le rendement diminue presque de moitié entre la première et la seconde année de culture, ce qui est attribué à une diminution de la fertilité du sol, qui n'est pas compensée par un apport d'engrais. Le rôle présumé de la jachère est de rétablir la fertilité du sol : la végétation naturelle qui a poussé pendant la jachère est enfouie dans le sol au moment du labour, pour servir de compost, et c'est plusieurs mois après cet enfouissement que commence la période de culture. Le modèle Faprom a été mis au point pour tenter de comprendre le rôle de la jachère dans ce processus.
Six espèces ont été simulées : une pionnière Rumex acetosella (introduite d'Europe), une espèce intermédiaire, la légumineuse Lupinus meridanus, trois arbustes Acaena elongata, Baccharis prunifolia et Hypericum laricifolium, et une espèce climacique endémique, Espeletia schultzii (composée en rosette géante emblématique du paramo).
2.3 Les mécanismes pris en compte dans le modèle
Pour chaque espèce, on calcule dans chaque strate horizontale la lumière absorbée, proportionnelle au produit de la surface foliaire locale par le coefficient d'absorption (qui dépend du port des feuilles). Puis on transforme cette lumière absorbée en photosynthèse connaissant la réponse photosynthétique de chaque espèce, mesurée en saison humide et en saison sèche, et on intègre sur la journée, en enlevant la part de la respiration de croissance pour obtenir l'entrée de carbone dans la plante (flux , Fig. 2). La respiration d'entretien est ensuite calculée pour chaque organe avec un coefficient qui est fonction de sa concentration en azote.
L'allocation du carbone à chaque organe se fait de manière à obtenir une biomasse par organe compatible avec les données observées sur le terrain, faute de disposer d'un modèle mécaniste d'allocation. La sénescence est modélisée par une équation du premier ordre : une fraction constante des tissus (différente cependant suivant les organes, les espèces et pour chacune des saisons) meurt à chaque pas de temps, assurant ainsi un taux de renouvellement de ces tissus. La reproduction est alimentée à chaque pas de temps (journée) par le flux de carbone qui est alloué aux parties reproductives, et une fois par an les graines sont dispersées. La banque de graines du sol est alimentée par cette production annuelle, par les graines en dormance et par la pluie de graines arrivant des parcelles voisines. Au début de la saison humide, une partie de la banque de graines germe et leur biomasse est attribuée directement aux organes de l'espèce considérée. C'est donc un traitement très simplifié du cycle reproductif.
Un modèle simple de matière organique a été utilisé pour simuler le cycle d'azote dans le sol. La litière est divisée en deux fractions, l'une décomposée rapidement et entièrement, l'autre enfouie comme matière organique du sol et décomposée également mais beaucoup plus lentement. Il en résulte un flux d'azote minéral en provenance de ces deux sources, et aussi des dépôts d'azote secs et humides. Cet azote minéral est en partie absorbé par les racines des plantes et en partie lixivié, en fonction du flux d'eau drainé et de la concentration en azote minéral dans la solution.
3 Sorties types du modèle dans la simulation de base
Ce modèle a été utilisé avec les valeurs par défaut des paramètres pour simuler la croissance en biomasse et en surface foliaire des six espèces représentées, pour une période de 20 ans au maximum. Globalement, la communauté atteint un indice foliaire (surface totale de feuilles par unité de surface de sol) maximal d'environ 2 après quatre ans de simulation environ, ce qui traduit la relative pauvreté du sol, avec un flux d'azote minéral de l'ordre de 4 g m−2 an−1 [3]. La biomasse totale croît rapidement pendant ces quatre premières années, puis beaucoup plus lentement, ce qui correspond à la croissance des organes pérennes (tiges, troncs et branches). La quantité d'azote mobilisée dans cette biomasse augmente aussi rapidement, atteignant 6 g m−2 après quatre ans, puis bien plus lentement jusqu'à 7 g m−2 à 12 ans. Il faut y ajouter 2 g m−2 pour la litière et environ autant pour la nécromasse sur pied, soit un total un peu supérieur à 10 g m−2.
Un point intéressant pour le rôle de la jachère est l'évolution du stock d'azote minéral du sol. Nous avons réalisé trois simulations en supposant, soit l'absence de toute végétation (sol nu), soit une communauté végétale sans légumineuse, soit encore une communauté complète avec légumineuse. Pour le sol nu, on observe une diminution rapide du stock d'azote du sol (de 4 à 5 g m−2 an−1), qui correspond au flux d'azote minéralisé et lixivié car non utilisé pour la croissance. Pour la végétation sans légumineuse, on observe une légère diminution les premières années, puis une stabilisation du stock un peu en dessous de la valeur de départ, ce qui signifie que les pertes par lixiviation sont entièrement compensées par les faibles apports atmosphériques. Enfin, pour la communauté avec légumineuse (simulation de base), l'azote du sol baisse légèrement puis augmente à nouveau et retrouve sa valeur initiale avant quatre ans, pour augmenter ensuite très légèrement jusqu'à 12 ans.
La croissance en biomasse des diverses espèces est représentée sur la Fig. 3 pour la simulation de base. Nous n'avons malheureusement pas de données mesurées avec lesquelles effectuer une comparaison quantitative. La raison en est l'hétérogénéité dans les conditions initiales réelles, qui entraîne des trajets successionnels différents et des végétations différentes pour un même âge d'abandon des parcelles. Toutefois, on retrouve qualitativement le comportement observé sur le terrain : prédominance de l'espèce pionnière Rumex la première année (sur le terrain, elle reste abondante pendant plusieurs années, plus que dans notre simulation), suivie de celle de l'espèce secondaire Lupinus (qui augmente à nouveau un peu de 9 à 12 ans), puis croissance des arbustes et de la rosette géante Espeletia. Dans cette simulation, la biomasse d'Espeletia et d'Hypericum diminue à partir de 6 ans, alors que celle d'Acaena et de Baccharis augmente de façon continue. Les six espèces demeurent à la fin des 12 années, mais en très faible biomasse pour Rumex [5,6].

Biomasses simulées des six espèces au cours de la jachère.
À ce stade, plusieurs questions intéressantes peuvent être posées :
- (a) quelles conditions sont nécessaires pour conserver une végétation plurispécifique ? On sait en effet que beaucoup de modèles ont des difficultés à simuler une co-dominance d'espèces et tendent vers la dominance d'une seule espèce ;
- (b) comment la végétation est-elle affectée par la disponibilité en ressources du milieu, en termes de fonctionnement global (production de biomasse, stockage d'azote) et de partage des ressources entre les différentes espèces ?
Nous allons présenter des résultats concernant la seconde question, et aborderons la première dans la discussion.
4 Variation de la production de biomasse en fonction des ressources disponibles
On entend par ressources en écologie les principaux facteurs susceptibles de modifier croissance et production des espèces étudiées. Dans une étude par simulation, il est évidemment facile de faire varier ces ressources : rayonnement solaire, alimentation en eau et en minéraux. Nous avons choisi de ne pas modifier le rayonnement solaire, qui varie peu pour un même lieu, et de nous concentrer sur les deux autres facteurs, en faisant varier le seul azote, puisque notre modèle ne simule pas les autres éléments minéraux. Cette fois, les simulations ont été conduites sur une période de 20 ans, avec toujours les valeurs par défaut des paramètres susceptibles de varier. Pour l'alimentation en eau, nous avons fait varier la durée de la saison humide de trois (traitement W1, le plus sec) à neuf mois (W5, le plus humide), le traitement moyen W3 correspondant à un climat réel moyen à Gavidia. L'alimentation en azote varie entre 1 g m−2 an−1 (traitement N1) et 8 g m−2 an−1 (traitement N8, pour lequel l'azote reste un facteur limitant), la situation réelle étant estimée entre N4 et N5.
La Fig. 4 présente la production totale de biomasse cumulée sur 20 ans de jachère pour ces divers traitements en eau et azote. Elle varie d'environ 250 à 1500 g m−2 an−1. Pour une faible disponibilité en azote (N1 à N3), la production augmente régulièrement avec l'augmentation de la durée de la saison humide de W1 à W5, ce qui est conforme à ce qu'on peut attendre. En revanche, pour un traitement en eau donné (W3 par exemple) la production diminue quand augmente la disponibilité en azote du sol, ce qui est tout à fait contraire à ce que suggère l'intuition. Cette diminution est peut-être la plus étonnante pour le traitement le plus humide W5, pour lequel la production diminue seulement pour N7 et surtout N8. Autrement dit, une bonne alimentation en eau et en azote peut aboutir à une végétation moins productive qu'une plus faible alimentation en azote. Comment expliquer cette simulation paradoxale ? Elle est sans aucun doute liée à la composition en espèces de la communauté, qui peut varier de façon importante avec le niveau de ressources, selon les paramètres affectés à chaque espèce.

Production de biomasse cumulée pendant 20 ans de jachère, en fonction de la disponibilité en eau et en azote.
Nous avons vu plus haut que chaque espèce est caractérisée par 48 paramètres, ce qui rend difficile la comparaison entre espèces faute de connaître quels sont les paramètres les plus importants. Pour tenter de hiérarchiser ces paramètres, nous avons construit un modèle simple de croissance pour une végétation monospécifique, et tenté de relier les trois paramètres de ce modèle simple aux 48 paramètres du modèle plus mécaniste Faprom.
5 Croissance simplifiée d'une végétation monospécifique
Nous avons d'abord simulé, avec le modèle complet, la croissance d'une végétation monospécifique. Lorsqu'on porte la biomasse en fonction du temps, on obtient une courbe en S très courante en biologie. Cette courbe a été décrite pour la première fois [7] à propos de la population humaine sous forme de l'équation logistique qui a été ensuite reprise [8]. Si x désigne la variable d'intérêt, cette équation s'écrit sous forme différentielle :
| (1) |
Elle est entièrement décrite par deux paramètres : le taux de croissance intrinsèque de la population (r), et la capacité de charge du milieu qui est la population maximale (ou biomasse maximale dans le cas d'une végétation) qui peut être supportée pour un niveau donné de ressources (en situation « d'équilibre »). Comment se fait-il qu'un modèle de croissance complexe arrive finalement à se comporter comme un modèle aussi simple ?
Le Tableau 1 présente les valeurs de r et de K ajustées pour les diverses espèces aux valeurs de biomasse simulées par le modèle complet (Faprom) avec les paramètres par défaut. On retrouve le classement écologique des espèces pionnières (r élevé, K faible) et de fin de succession (r faible, K élevé), avec deux intermédiaires, dont une (Acaena) n'est pas différentiable des espèces de fin de succession.
Caractéristiques de croissance et statut de succession des espèces étudiées. r et K sont obtenus par ajustement logistique sur les sorties simulées par Faprom pour les végétations monospécifiques ; les statuts de succession sont établis sur les études de Sarmiento et al. (2003) : P, espèce pionnière, I, espèce intermédiaire, F, espèce de fin de succession. R = Rumex acetosella, L = Lupinus meridanus, A = Acaena elongata, E = Espeletia schultzii, B = Baccharis prunifolia, H = Hypericum laricifolium
| Site | Paramo de Gavidia (Venezuela) | |||||
| Espèce | R | L | A | E | B | H |
| r (g g−1a−1) | 6,9 | 2,4 | 1,3 | 1,9 | 1,1 | 1,3 |
| K (g m−2) | 195 | 579 | 786 | 636 | 773 | 830 |
| Statut de succession | P | I | I | F | F | F |
Comment varient r et K en fonction des paramètres de Faprom ? Nous avons réalisé une analyse de sensibilité et en présentons les résultats dans le Tableau 2, en séparant les espèces non légumineuses (qui sont moyennées, car leurs réponses sont très semblables) et celles de la légumineuse. L'élasticité est la variation relative (de K ou r) rapportée à la variation relative du paramètre testé. Elle a été obtenue en moyennant les valeurs d'élasticité obtenues pour six variations relatives de chaque paramètre (−15%, −10%, −5%, 5%, 10%, 15%). Pour les espèces non légumineuses, r est relié négativement à la masse surfacique des feuilles (LMA) et positivement aux taux de photosynthèse (P) et de sénescence (S), qui sont liés puisqu'on arrive à une biomasse maximale. Pour la légumineuse, r est aussi fortement relié négativement à la masse surfacique et positivement à la photosynthèse, mais négativement relié à la mortalité et à la teneur en azote des feuilles (qui augmente leur respiration, cet effet n'existant pas pour les espèces non légumineuses, qui sont toujours fortement limitées en azote).
Influence des paramètres du modèle Faprom sur les paramètres r et K de l'équation logistique ajustée sur la sortie de Faprom. L'élasticité est la variation relative (de r ou K) sur la variation relative du paramètre de Faprom 8. et sont les paramètres de respiration de croissance et d'entretien, LMA est la masse surfacique des feuilles, P, S et N sont des facteurs d'échelle pour la photosynthèse, la sénescence et la teneur en azote, respectivement
| Espèce | Élasticité de r pour divers paramètres | |||||
| LMA | P | S | N | |||
| Non-légumineuse | −0,10 | −0,07 | −0,60 | 0,38 | 0,55 | −0,09 |
| Lupinus | −0,50 | −0,50 | −2,45 | 2,70 | −0,8 | −0,51 |
| Espèce | Élasticité de K pour divers paramètres | |||||
| LMA | P | S | N | |||
| Non-légumineuse | −0,05 | −0,15 | −0,18 | 0,35 | −0,98 | −1,05 |
| Lupinus | −0,54 | −0,80 | −1,05 | 2,03 | −1,15 | −0,82 |
Pour ce qui concerne la biomasse maximale K, elle est reliée négativement à la sénescence et à la teneur en azote pour les espèces non légumineuses ; pour la légumineuse, K est fortement liée positivement à la photosynthèse et négativement aux autres paramètres : sénescence, teneur en azote, masse surfacique, taux de respiration.
Les paramètres r et K définis sur les simulations de végétations monospécifiques ne sont pas sensibles à la hauteur maximale, puisqu'il n'y a pas de compétition entre espèces pour la lumière ; ils ne sont pas non plus sensibles à la longueur de racine par unité de masse, puisqu'il n'y a pas de compétition interspécifique pour l'azote. En effet, le modèle utilise la hauteur maximale uniquement pour répartir la surface foliaire dans les strates de couvert. Pour une végétation monospécifique, la lumière absorbée dépend uniquement de la surface foliaire totale et non de sa distribution dans les strates. En revanche, dans un mélange d'espèces, le modèle est sensible à la définition des hauteurs maximales et plus particulièrement des hauteurs relatives entre les différentes espèces. En ce qui concerne l'absorption d'azote, le modèle ne simule pas la capacité absolue d'absorption des différentes espèces, mais uniquement leur capacité relative. Une espèce seule peut (dans le modèle) absorber tout l'azote minéral disponible, quelle que soit sa biomasse de racines.
L'équation logistique présente l'inconvénient de ne pas séparer les processus antagonistes d'assimilation et de mortalité simulés dans le modèle Faprom. Pour cela Martineau [3] a exprimé ces processus de façon simplifiée par rapport à Faprom, mais plus explicite par rapport au modèle logistique, en séparant les gains de carbone (issus de la photosynthèse) et les pertes (par mortalité et respiration d'entretien) dans l'équation suivante :
| (2) |
Cette équation, appelée « pseudo-logistique », fournit comme la logistique une courbe en S, mais elle possède trois paramètres au lieu de deux, ce qui permet un ajustement plus fin des données. Ces paramètres sont A (g m−2 j−1), taux maximal d'assimilation, a (g g−1 j−1), taux d'assimilation par unité de biomasse, et q (j−1) taux de mortalité (et de respiration d'entretien). x est la biomasse en g m−2. Martineau [3] a montré que cette équation est formellement identique à celle proposée par Smith [9] sous la forme :
| (3) |
On voit que l'équation de Smith dérive de la logistique en rajoutant au dénominateur un terme contenant le paramètre q de mortalité, correspondant au turn-over de la biomasse.
Devant la simplicité de ces modèles comparée à la complexité du modèle mécaniste Faprom, on peut se demander quel est l'intérêt de ce dernier. La réponse est simple. Les modèles simplifiés peuvent seulement rendre compte de la croissance dans des conditions de milieu définies (lumière, alimentation en eau et en azote). Ils ne peuvent être utilisés de façon plus générale (et bien sûr dans un modèle où les espèces sont en compétition) que si on connaît la façon dont leurs paramètres dépendent de ces facteurs du milieu. Cette dépendance est automatique dans Faprom 8, puisqu'elle est inscrite dans les équations du modèle. Elle est illustrée pour le modèle simplifié dans la Fig. 5 pour la lumière et dans la Fig. 6 pour l'azote. On a supposé que le paramètre q de mortalité de l'Éq. (2) ne variait pas avec la lumière ou l'azote. Il en résulte que le taux absolu de mortalité B (égal à qx, où x est la biomasse) est représenté par une droite en gras dans les deux figures. L'ombrage réduit à la fois la pente à l'origine et le plateau de la courbe d'assimilation (Fig. 5, courbes A, Ombr+ et Ombr++), alors que la carence en azote réduit seulement le plateau (assimilation maximale, Fig. 6, courbes A, N– et N– –). La biomasse à l'équilibre, qui correspond à l'abscisse du point d'intersection entre la courbe de croissance et la droite de mortalité, est donc affectée de façon différente par la lumière et par l'azote.

Représentations des composantes de la croissance dans la courbe pseudo-logistique (Éq. (2)), en fonction de la biomasse x (gC m−2). A (gC m−2 j−1) représente l'assimilation de carbone en éclairement naturel, et Ombr+ et Ombr++ sa valeur réduite par deux niveaux d'ombrage. B représente les pertes par mortalité, produit d'un taux relatif constant q par la biomasse x.

Représentations des composantes de la croissance dans la courbe pseudo-logistique (équation (2)), en fonction de la biomasse x (gC m−2). A (gC m−2 j−1) représente l'assimilation de carbone en alimentation azotée normale, et N– et N– – sa valeur réduite par deux niveaux de carence azotée. B représente les pertes par mortalité, produit d'un taux relatif constant q par la biomasse x.
6 Discussion et interprétation
Les résultats de la Fig. 4 ont montré une diminution de la production globale de la végétation en réponse à une augmentation de la disponibilité en azote. Ils peuvent maintenant être interprétés connaissant les caractéristiques des diverses espèces simulées et leur sensibilité à la compétition. La Fig. 7 présente les variations d'indice foliaire de chaque espèce pour diverses disponibilités en eau et en azote. On a utilisé l'indice foliaire plutôt que la biomasse dans cette figure pour pouvoir plus facilement comparer les espèces, le recouvrement étant lié à l'indice foliaire plutôt qu'à la biomasse. Pour des raisons de lisibilité, contrairement à la Fig. 4, on n'a conservé que trois valeurs de chaque traitement, les valeurs extrêmes et une valeur intermédiaire ; l'ensemble des simulations est disponible dans [3]. L'échelle des ordonnées a été ajustée aussi pour des raisons de lisibilité. Les valeurs intermédiaires W3 et N5 correspondent à la simulation de base.

Évolution de l'indice foliaire des six espèces, simulée pendant 20 ans de jachère, pour diverses alimentations en eau et en azote. Symboles des espèces : A = Acaena elongata, B = Baccharis prunifolia, E = Espeletia schultzii, H = Hypericum laricifolium, L = Lupinus meridanus, R = Rumex acetosella. W1 à W5 représentent des niveaux croissants d'alimentation en eau (saison sèche variant de 9 mois en W1 à 3 mois en W5), N1 à N8 des niveaux croissants d'alimentation azotée (de 1 à 8 gN m−2 an−1).
Lorsqu'il y a très peu d'azote (traitement N1), c'est la légumineuse qui domine, comme on peut s'y attendre. Mais comme elle est assez sensible au déficit hydrique, d'autres espèces arbustives moins sensibles sont aussi présentes dans le traitement W1 : Baccharis et Hypericum.
Lorsqu'il y a beaucoup d'azote (N8), diverses espèces dominent selon la disponibilité en eau : Espeletia dans le traitement sec W1, puis Acaena en W3, puis la légumineuse Lupinus avec co-dominance d'Acaena dans le traitement le plus humide W5.
Dans le traitement intermédiaire (N5), Baccharis et Hypericum dominent en W1 (avec une apparition transitoire d'Espeletia les premières années). En W3, Acaena augmente rapidement, ainsi que Lupinus les premières années, et en W5 Lupinus domine avec Acaena augmentant lentement.
Il est clair d'après la Fig. 7 qu'on observe le plus grand nombre d'espèces dans la situation intermédiaire (N5, W3) lorsqu'à la fois l'eau et l'azote sont limitants. Pour comprendre la réduction de la production globale observée sur la Fig. 4 lorsque augmente la disponibilité en azote, il faut observer la Fig. 7 par colonne en descendant (augmentation de l'alimentation en azote pour une même alimentation en eau). Pour les traitements W1 et W3, Lupinus, qui domine en N1, est entièrement remplacé en N8 par une autre espèce : Espeletia (qui résiste bien à la sécheresse) en W1, et Acaena en W3. Espeletia comme Acaena sont plus hautes que Lupinus et lui portent ombrage, malgré une croissance initiale plus faible. Pour le traitement le plus humide W5, Lupinus reste dominant mais Acaena est co-dominant. Dans chaque cas, Lupinus se fait donc remplacer par une autre espèce à photosynthèse et croissance plus faibles. C'est donc ce remplacement d'une espèce productive par une espèce qui l'est moins qui entraîne la baisse de la production globale observée dans la Fig. 4.
7 Conclusion
Nous avons montré, à l'aide d'un modèle mécaniste simulant la compétition entre six espèces de statut écologique différent, que la production végétale pouvait diminuer en réponse à une augmentation des ressources du milieu. Cette diminution provient d'un remplacement d'espèces : dans notre cas, la légumineuse, qui domine lorsque le sol est carencé en azote, est remplacée par des espèces moins productives lorsque le sol s'enrichit. Cette situation pourrait avoir une portée générale. Ainsi lorsqu'on étudie la réponse d'espèces isolées à une augmentation de teneur en CO2, on trouve habituellement une forte stimulation, avec cependant des différences entre espèces. On en déduit, en général, que la production végétale doit augmenter en réponse à cette augmentation du CO2. Dans les conditions naturelles, les espèces sont en compétition pour d'autres ressources que le CO2 : lumière, eau, minéraux. Ces autres facteurs agissent en interaction avec le CO2, et il n'est donc pas possible d'extrapoler simplement à une végétation plurispécifique les résultats obtenus sur quelques espèces cultivées isolément. Ce raisonnement pourrait expliquer la plus faible stimulation de croissance par le CO2 des communautés herbacées par rapport aux forêts qui, au moins en Europe, présentent un nombre d'espèces beaucoup plus limité [10]. Un autre exemple est, dans les régions tropicales à saison sèche, la progression des arbres sur les herbes dans les savanes. Une explication de cette progression est que la croissance des arbres ou des arbustes (espèces en C3) est plus fortement stimulée par l'augmentation du CO2 atmosphérique que celle des herbes en C4 [11]. La réduction de l'ouverture stomatique en CO2 élevé entraîne une meilleure disponibilité en eau du sol, qui profite plus aux arbres qu'aux herbes. Il n'est pas impossible que le résultat net soit une diminution de la production de ces savanes, les arbres étant globalement moins productifs que les herbes.
Remerciements
Ce travail a été réalisé dans le cadre d'une thèse financée par le ministère français de l'Éducation nationale, de l'Enseignement supérieur et de la Recherche, au sein de l'École doctorale « Sciences du Végétal », à Orsay. Il a été soutenu par la Communauté européenne dans le cadre du programme Inco Tropandes [2]. Les auteurs remercient particulièrement la coordinatrice, le Pr. Tarsy Carballas (université de Saint-Jacques-de-Compostelle), la responsable du programme au Venezuela, le Pr. Lina Sarmiento (Universidad de Los Andes, Merida, Venezuela), pour les données météorologiques, et le Dr Pierre Bottner (CNRS, Montpellier), pour les données concernant la fourniture d'azote minéral.