


 CC-BY 4.0
CC-BY 4.0
1. Introduction
The Arum genus is composed of 29 described species [1, 2, 3] but estimated to be comprised of 43 species [4]. Regardless of the genus diversity, one stable subdivision within the genus, based on morpho/ecological characters and molecular data, consistently groups all Arum species separately from Arum pictum. A. pictum is the only species of the subgenus Gymnomesium (L.f.) Schott, which appears to be sister to all other Arum species [1, 2, 5, 6]. Thus, A. pictum appears to be a unique taxon within the genus Arum, endemic to West Mediterranean islands: Corsica, Sardinia, Tuscan Archipelago (Montecristo) and Balearic Islands [1]. This species presents some distinguishing characters such as (1) autumn flowering (versus spring for all other species) and (2) leaves produced after or simultaneously with inflorescences (versus before for all other species, [2]). For all of the above-cited reasons, it has been proposed to elevate the subgenus Gymnomesium to the rank of a monospecific genus [4]. Since a proper taxonomical synonymy has not yet been published, the name Arum pictum will be used in this manuscript.
The inflorescence of A. pictum consists of a spadix (e.g. reproductive spike) and a spathe (e.g. modified bract). The anthesis lasts two days and is protogynous, preventing spontaneous self-pollination [7]. The spathe, which completely encloses the spadix during floral development, opens in its upper half during anthesis to reveal the sterile appendix of the apical part of the spadix. This appendix produces heat and releases scent for pollinator attraction. The basal half of the spathe forms a pollination chamber around the basal part of the spadix, which bears unisexual flowers, stamens and ovaries. Attracted insects are trapped on the first day of anthesis during the female stage when they slip and fall into the floral chamber, and eventually pollinate the female flowers. On the next morning, during the male stage, they are dusted with pollen before being able to leave the pollination chamber. After pollination in autumn, red berry-like fruits develop throughout the winter as an infructescence that mature by the following spring [7].
The deceptive pollination strategy of A. pictum has been studied more than a decade ago in one Corsican population [7]. In summary, the anthesis cycle of A. pictum occurred during the day, attracting mainly sphaerocerid flies (Brachycera) and staphylinid beetles early in the morning and late in the afternoon of the first day. The pollen was released from the anthers early in the morning of the second day before insects depart. A. pictum’s thermogenic cycle matched the anthesis cycle and the pollination events with an original and unique, bimodal temperature pattern of the appendix (early morning ∼8:00 am and late afternoon ∼07:30 pm), contrary to the unimodal pattern found in all other studied Arum species [7, 8]. As mentioned earlier, the main pollinators of Arum pictum were sphaerocerid flies (Brachycera) and staphylinid beetles and not psychodid, chironomid or ceratopogonid flies (all nematoceran) known to be the main pollinators of Arum species in West Europe [8, 9, 10, 11, 12, 13]. Most of the Coproica (Sphaeroceridae) flies captured by A. pictum inflorescences were females (more than 70%) suggesting that the floral scent of A. pictum may have sex-specific properties and/or may mimic the fly oviposition sites [7]. A. pictum is characterised by a dung odour close to horse/donkey dung, described as pungent [1]. The floral scent was described as been dominated by 5 volatile compounds namely: benzyl alcohol, indole, dihydroocimene, ocimene and skatole [7]. The comparative study of the volatiles compounds (VOCs) emitted by four different types of dung showed that the scent of the horse dung was dominated by p-cresol, β-citronellene, and 4-methyl-3-heptanone [14]. Other notables VOCs were dihydrolimonene, cis-pinane, indole, skatole, and various alkanes [14].
The aim of this paper is to document some regional aspects of the pollination ecology of Arum pictum across the island of Sardinia (Italy) in comparison to Corsica. We measured the floral thermogenic patterns, collected the floral volatiles during the most scented phase of the anthesis according to human nose, and the insects trapped within pollination chambers of inflorescences from several populations.
2. Materials and methods
2.1. Study species and populations
Arum pictum L. fil. is a Tyrrhenian islands endemic, found on Sardinia, Corsica, Mallorca, Menorca and Montecristo in the Tuscan Archipelago [15, 16], including some minor islets surrounding the large islands. In Sardinia, it is distributed in all sectors of the islands [16], usually below 1000 masl, but sometimes above [17, 18]. The plant thrives on every kind of soil, but prefers shady or semi-shady habitats under the canopy of evergreen trees and shrubs [19] in the thermo- and meso-Mediterranean bioclimatic belts [20]. This species is not threatened in Italy and its conservation status is considered as LC [21].
The pollination ecology of Arum pictum was studied in October 2018 in three populations from northern Sardinia (Italy). The first two populations were located in north-western Sardinia, namely: near the lake Baratz (Sassari, GPS coordinates: 40.676729, 8.225554) and Gutierrez (Alghero, GPS coordinates: 40.625107, 8.273025) at almost sea level (24 masl), in the thermo-Mediterranean dry bioclimatic belt, the first on aeolian sand, the second on Mesozoic limestone. Both populations grew in Pinus pinea and P. halepensis plantations, but the first is characterized by Juniperus phoenicea vegetation, whereas the second by Olea europaea vegetation. The third population was located in north-eastern Sardinia at Diga del Liscia (Sant’Antonio di Gallura-Luras, GPS coordinates: 41.001154, 9.144749, named Diga afterwards) at an elevation of 242 masl, corresponding to a meso-Mediterranean dry bioclimatic belt. This population grew on acidic soil (on schists and granites) in an open wood of Quercus suber. In autumn 2018, these three Arum populations were large with more than 100 individual plants. For a broader geographical comparison, one south-west Corsican population was also studied in October 2018, located in Coti-Chiavari (GPS coordinates: 41.793473, 8.769976, named Coti afterwards) on the south bank of the Ajaccio Gulf in a former penitentiary area. This population, large of ∼60 individual plants, grows in an open wood of Quercus ilex.
2.2. Thermogenesis and anthesis
Thermogenesis in A. pictum was studied by measuring the temperatures of 8 inflorescences belonging to two different Sardinian populations (Baratz in the west and Diga in the east). During the entire anthesis, temperatures were recorded every 10 min from two different zones of the spadix (the appendix and the male flowers) with simultaneous measurements of ambient air, using type-K probes connected to Digi-Sense® DualLogR® thermocouple thermometers (Testo model 175T3®). The female flowers (e.g. gynoecia) are known to be non-thermogenic [7] and were thus not recorded. The two probes recording the spadix temperatures were inserted about 5 mm deep into either the appendix or the stamen zone. Ambient air temperature was recorded near the studied inflorescence in the shade (e.g. under an Arum leaf). Twice per day (i.e. in the morning and in the afternoon) the monitored inflorescences were observed for insect presence and floral observations.
2.3. Floral scent analyses
Floral volatiles were collected during the morning between 09:00–10:30 am in the four studied populations from Coti (N = 9 inflorescences) between October 4th and 7th, Gutierrez (N = 3) on October 13th, Baratz (N = 2) on October 14th, and Diga (N = 4) on October 19th. Floral scent samples were collected in situ from scented inflorescences that were enclosed within Nalophan (NA colourless, calibre 90, ETS Charles-Frères, Saint-Etienne, France) bags opened in their upper part to create a headspace. The emitted volatile organic compounds (VOCs) were adsorbed by Solid Phase Micro Extraction (SPME) Fibers (StableFlexTM SPME Fiber, 65 μm PDMS-DVB Coating, Supelco, Darmstadt, Germany) for 20 min, then stored at 5 °C until analysis by thermal desorption. To identify possible ambient contaminants, control samples were simultaneously collected from an empty bag for each scent sample, following the same protocols described above.
The trapped volatiles were analysed by combined gas chromatography-mass spectrometry (GC-MS). The SPME fibers were manually injected and desorbed into the split/splitless injector of a Clarus 580 Perkin-Elmer directly coupled to a Clarus SQ8S Perkin Elmer TurboMass detector (quadrupole). The gas chromatographer was equipped with two fused-silica capillary columns (50 m × 0.22 mm, film thickness 0.25 μm), a non-polar BP-1 (polydimethylsiloxane) and a polar BP-20 (polyethylene glycol). The oven temperature was programmed from 60 to 220 °C at 2 °C⋅min−1 and then held isothermal at 220 °C for 20 min; injector temperature at 270 °C; carrier gas was helium (0.8 mL⋅min−1); split ratio of 1:15; ionisation energy at 70 eV. The electron ionisation (EI) mass spectra were acquired over the mass range 40–350 Da. The relative proportions of the constituents were expressed as percentages obtained by peak-area normalisation, without using correcting factors. Kovats retention indices (RI) were determined relative to the retention times of a series of n-alkanes with linear interpolation (Target Compounds software from Perkin Elmer). Compounds were tentatively identified by comparing their mass spectra and retention times to those of authentic standards and from the reference libraries Adams, NIST20, Perkin FF2013, Wiley FFNSC Library 2nd Edition and Wiley Registry™ 8th Edition.
2.4. Isomyocorene NMR identification
In order to confirm mass identification of the main unidentified floral VOCs, floral scent samples were collected in situ from four inflorescences during the peak of thermogenic activity and floral scent emission on the female phase of anthesis. The inflorescences were enclosed within Nalophan (NA colourless, calibre 90, ETS Charles-Frères, Saint-Etienne, France) bags opened in their upper part to create a headspace. Sampling was conducted by dynamic headspace, scented air was drawn for 30 min through adsorbent traps connected with silicone tubing to a membrane pump (ASF Thomas Inc, Memmingen, Germany) that worked at a constant flow of 250 mL⋅min−1. The adsorbent traps (10 cm long, 0.5 i.d.), made out of silanised glass cartridges, were filled with a 1:1 mixture of Tenax™ TA (50 mg, 80/100, Macherey-Nagel 706318, Düren, Germany) and Carbopack™ X (20/40, Supelco 1-0435), fixed in each tube with silanised glass wool plugs.
After collection, the four traps were eluted with deuterated chloroform (150 μL CDCl3) and pooled together to attain a sufficient quantity for the NMR analysis. Nuclear Magnetic Resonance spectra were recorded on a Bruker AVANCE 400 Fourier Transform spectrometer (Wissembourg, France) operating at 400.132 MHz for 1H and 100.623 MHz for 13C, equipped with a 5 mm probe, in CDCl3, with all shifts referenced to internal tetramethylsilane (TMS). The 1H-NMR spectrum was recorded with the following parameters: pulse width (PW) 4.3 μs; relaxation delay 1 s, acquisition time: 2.6 s for 32 K data table with a spectral width (SW) of 6000 Hz; accumulation scans: 32. The 13C-NMR spectrum was recorded with the following parameters: PW = 4 μs (flip angle 45°); acquisition time: 2.7 s for 128 K data table with a spectral width of 25,000 Hz (250 ppm); accumulated scans: 512. For the 2D spectra, sequences from Bruker Topspin™ (Bruker, Wissembourg, France) library (DEPT, COSY, HMBC) and gradient-enhanced sequences were used.
2.5. Visiting entomofauna
Arthropods attracted to the inflorescences and temporarily trapped in the pollination chamber were collected in ethanol 60% with an aspirator in three populations Coti (N = 26 inflorescences), Baratz (N = 30) and Diga (N = 8). Arthropods per inflorescence were counted and identified at the order rank (and family level for Staphylinidae and Braconidae). Based on known pollinators in the genus Arum, we grouped non-insect arthropods plus scarce insects (Formicidae, Blattoidea, Lepidoptera, Hemiptera) in the category “Others” [8].
3. Results and discussion
3.1. Thermogenesis and anthesis
The timing and intensity of the temperature peaks varied among inflorescences due to asynchronous flowering, inflorescence size, and physiology differences but the daily thermogenic pattern (e.g. sequence) was the same in individuals and populations. The appendix and the male flowers appeared to be responsible for heat production in inflorescences of Arum pictum but the two regions of the spadix were not synchronised with each other resulting in a complex thermogenic pattern (Figure 1). The first temperature rise, caused by the male flowers, was recorded the night before the opening of the spathe from 6:30 pm until noon the next day (Figure 1—sta1). Between 09:00–10:00 am, the temperature of the flowers reached a maximum of 23.2 °C, on average, 7 °C warmer than ambient air (Table 1). Later, on the same day, a second thermogenic phase occurred. The stamen temperature increased again between 3:30 and 9:30 pm (Figure 1—sta2) coinciding with the spathe opening (upper half). The temperature of the male flowers reached a maximum of 19–24 °C between 6:00–8:00 pm, which was, on average, 3.5 °C warmer than ambient air (Table 1). In the same evening, the stamen temperature increased again and with greater intensity between 9:30 pm and 3:30 pm on the following day (Figure 1—sta3). The temperature of the flowers reached a maximum of 22–30 °C between 01:30–04:30 am, which was, on average, 11.4 °C warmer than ambient air (Table 1). The stigmas looked wet with turgescent papillae (e.g. receptive) in early morning. The appendix started heating up at about the same time as the male flowers reached maximum temperatures (sta3), of 28–33 °C between 06:10–09:20 am (Figure 1—app1), corresponding to a maximum temperature difference of 9–20 °C above ambient air (Table 1). Insects were attracted to the scented inflorescences and eventually slipped or walked down inside the pollination chamber. Note that the stamen’s temperature increased again during the appendix thermogenic peak to a tissue temperature of 30–31 °C at 08:20–08:35 am representing a maximum temperature difference of 8.5–9.5 °C above ambient air (Figure 1). Afterwards, both stamen and appendix temperatures declined within two hours to reach ambient temperature at around noon. A third thermogenic phase began at the end of this first day of flowering: the appendix started heating up at about 1:40–4:00 pm (Figure 1—app2), while the male flowers started heating up at around 3:00–6:00 pm (Figure 1—sta4). Insects could also be attracted in the afternoon during this second scented phase but in somewhat lower numbers. Appendix temperature quickly reached a maximum temperature around 21–30 °C between 5:00–7:30 pm (4.7–12.7 °C above ambient air). After 6:00–7:00 pm, the appendix temperature decreased gradually until reaching ambient level between 9:20 pm and midnight (Figure 1). The male flowers’ temperature was still increasing and reached a maximum temperature of 21–26 °C (3.75–9 °C above ambient air, Table 1) around 00:30–03:00 am on the morning of the second day (Figure 1—sta4). Later in the morning, the pollen was released from the mature stamens and the trapped insects were able to leave the pollination chamber. Afterwards, the temperature declined, returning abruptly to ambient level between 10:30 am and noon.
Temperature difference curves from one inflorescence from Baratz representative of the typical thermogenic pattern in Arum pictum: between the appendix and ambient air (black line) and between the male flowers and ambient air (brown line). The grey boxes indicate the night period. The temperature peaks are numbered by chronological order for both the stamens “sta” and the appendix “app”.
Thermogenic data of Sardinian Arum pictum recorded from 10 inflorescences
| Temperature peaks | Anthesis time (days) | Mean duration (h) (min–max) | Temp. diff. max (°C) (min–max) |
|---|---|---|---|
| Stamens 1 | D-2/D-1 | >17h25a | 6.9 (6.7–7) |
| Stamens 2 | D-1 | 6h10 (4h30–8h00) | 3.5 (2.5–5.1) |
| Stamens 3 | D-1/D | 15h30 (14h20–17h30) | 11.4 (7.1–12.6) |
| Appendix 1 | D | 10h56 (8h50–12h30) | 14.6 (9.3–20.2) |
| Appendix 2 | D | 8h32 (6h20–10h10) | 6.7 (4.7–12.7) |
| Stamens 4 | D/D+1 | 17h56 (17h00–19h40) | 7.5 (3.7–9) |
For each temperature peak: corresponding day(s) during the anthesis, mean duration of the thermogenic event (range), and mean maximum temperature difference (range). D: pollination day.
a Temperatures were recorded after the temperature increase started.
The floral thermogenic pattern of Arum pictum individuals in Sardinia appeared to be consistent with the thermogenic pattern obtained from Corsican plants (see Figures 1B and 1C in [7]). A complex thermogenic pattern resulting from non-synchronised heat productions by two different regions of the spadix (e.g. the appendix and the male flowers) appears to be the rule in the genus Arumsensu lato [8] as it has been previously documented in Arum maculatum [22], A. italicum [23] and A. concinnatum [24]. Floral heat production in Arum results from an alternative mitochondrial respiration, the cyanide-insensitive pathway, catalysed by the alternative oxidase (AOX) resulting in heat dissipation instead of ATP production [24, 25]. The thermogenic pattern should be considered as a floral trait such as colour, shape or scent as the floral temperature peaks are synchronised with the main anthesis events in relation to the pollination process. The timing of insect attraction (e.g. scent emission and appendix temperature peak) occurred during the early morning and at the end of the afternoon. Morning temperature floral peak has been reported in some Arum species from East Mediterranean such as A. dioscoridis, A. creticum and A. palaestinum [24]; whereas in other species such as A. italicum, A. maculatum, A. rupicola and A. jacquemontii the temperature appendix peak has been recorded during the first evening or night of anthesis (review in [8]). The main originality of the thermogenic pattern in Arum pictum, in contrast with all other documented Arum species, is that the appendix is able to increase its temperature twice within the same day in a manner similar to fertile male flowers [7, 8, 23, 24]. This double temperature increase may be related to maximise day insect attraction in autumn, a potentially very temperamental season.
3.2. Isomyocorene identification
The major VOC was identified as 2,6-dimethyl-1,3,7-octatriene (Figure 2, supplementary materiel) by comparison of the proton and carbon chemical shift values with those reported in the literature [26].
Structure of the 2,6-dimethyl-1,3E,7-octatriene also known as isomyocorene. Right. Corresponding mass spectra.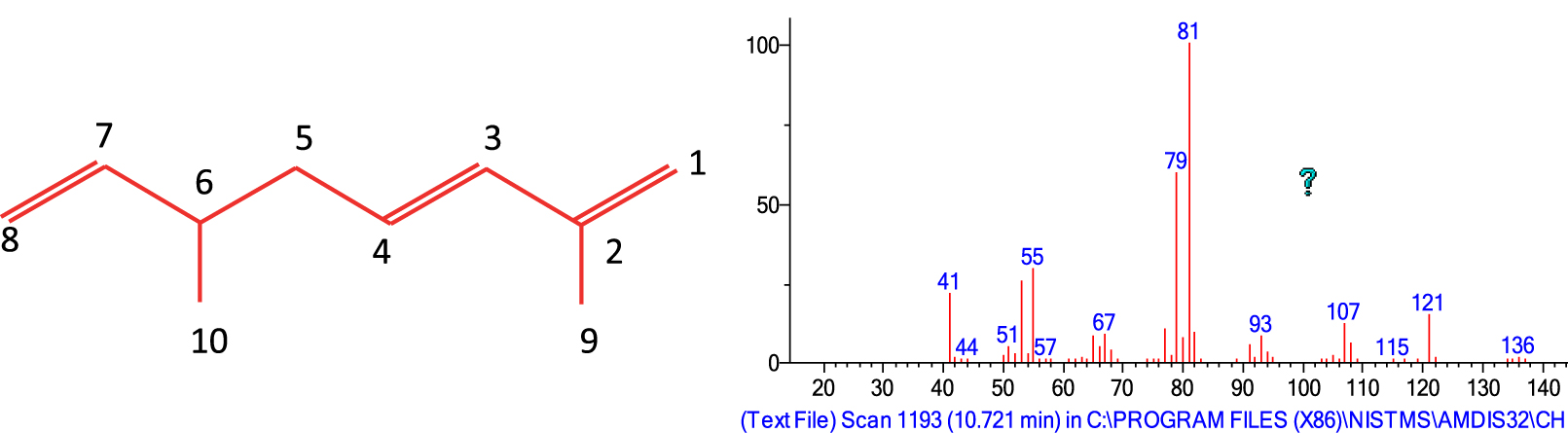
1H NMR (CDCl3, 400 MHz): 6.14 (d, H3); 5.77 (ddd, H7); 5.62 (dt, H4); 4.94–5.00 (m, 2H, H8); 4.87 (bs, 2H, H1); 2.22 (m, H6); 2.11 (m, 2H, H5); 1.83 (s, Me, H9); 1.00 (d, Me, H10).
13C NMR (CDCl3, 100 MHz): 144.09 (C7); 142.16 (C2); 134.14 (C3); 128.90 (C4); 114.40 (C1); 112.60 (C8); 39.90 (C5); 37.78 (C6); 19.50 (C9); 18.72 (C10).
EI-MS 70 eV, m/z (rel.int): 136 (1), 121 (12), 107 (12), 79 (57), 77 (10), 81 (100), 55 (27), 53 (22), 41 (22), 39 (16), 40 (11).
Kovats retention Indices: non-polar (BP-1): 960; polar (BP-20): 1119.
To the best of our knowledge, 2,6-dimethylocta-1,3,7-triene, also known as isomyocorene or alpha-menthrene, was obtained by palladium-catalysed telomerisation of isoprene [26]. It has been recently reported as a minor VOC emitted by scentless flowers of Tillsandia bandensis [27]. However, we have a doubt concerning the assignations of isomyocorene and cosmene in [27]. The authors reported isomyocorene with a Kovats retention index value of 1017 and cosmene with a Kovats retention index value of 968 (DB-5MS). Indeed, the Kovats retention index value of 1017 appears more adequate for cosmene isomers which are reported to elute after ocimenes (3,7-dimethyl-octatrienes) [28]. On the other hand, the Kovats retention index assigned to cosmene (RI: 968 DB-5MS) appears to be more appropriate for isomyocorene according to our data (RI: 960; BP-1). Then either these two compounds have been reversed or they have been misidentified [27].
3.3. Floral scent composition
In all samples, isomyocorene (see above) was the main floral VOC representing between 43% and 87% of the floral scent composition. Its corresponding alcohol 2,6-dimethyl-1,3,7-octatrien-6-ol (synonym hotrienol) was tentatively identified in small amounts in a few samples.
The floral scent of the Corsican population of Coti was mainly composed of seven VOCs representing 87–99% of the total blend (Table 2), namely: isomyocorene (average 72%), p-cresol (8%), β-citronellene (6%), indole and skatole (3% each), unknown1 and unknown4 (2% each). The floral scent of the west Sardinian population of Baratz was also mainly composed of seven VOCs representing 98–100% of the total blend (Table 2), namely: isomyocorene (average 62%), p-cresol (23%), β-citronellene (4%), 3-methylbutanoic acid (4%), skatole (3%), indole (1%), and unknown1 (1%). The floral scent of the west Sardinian population of Gutierrez was also mainly composed of seven VOCs representing 98–100% of the total blend (Table 2), namely: isomyocorene (average 71%), p-cresol (17%), 3-methylbutanoic acid (3%), β-citronellene and unknown3 (2% each), skatole and 3-methyl-1-butanol (1.2% each). Finally, the floral scent of the east Sardinian population of Diga was composed of only four main VOCs representing 96–99% of the total blend (Table 2), namely: isomyocorene (average 82%), β-citronellene (9%), p-cresol (6%), and unknown1 (1%).
Floral scent composition as blend relative proportion (%): mean, standard deviation, minimum and maximum from four populations (Corsica: Coti; Sardinia: Baratz, Gutierrez, Diga — see materials and methods for full details) of Arum pictum
| RI polar | RI non-polar | Name | Coti (N = 9) | Baratz (N = 2) | Gutierrez (N = 3) | Diga (N = 4) | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| mean ± stdev | min–max | mean ± stdev | min–max | mean ± stdev | min–max | mean ± stdev | min–max | |||
| 1030 | 937 | β-citronellene | 6.3 ±3.3 | 3.4–12 | 4.4 ±4.5 | 1.2–7.6 | 2.4 ±2.4 | 0.5–5.2 | 9 ±2.7 | 5.4–11.1 |
| 1119 | 960 | Isomyocorene | 71.5 ±11.2 | 57.5–87.4 | 62 ±26.5 | 43.2–80.7 | 71.3 ±12.2 | 60.1–84.3 | 82 ±2.1 | 79.2–83.9 |
| 1156 | 979 | Myrcene | 0.2 ± 0.2 | 0–0.4 | 0.1 ± 0.2 | 0–0.2 | 0 | 0 | 0.1 ± 0.1 | 0–0.2 |
| 1168 | 984 | Unknown1b | 1.8 ±0.3 | 1.3–2.3 | 1.2 ±1.7 | 0–2.4 | 0.2 ± 0.3 | 0–0.6 | 1.4 ±0.7 | 0.6–2.1 |
| 1181 | 997 | Unknown2b | 0.9 ± 0.4 | 0.4–1.6 | 0.5 ± 0.7 | 0–1 | 0 | 0 | 0.2 ± 0.3 | 0–0.5 |
| 1205 | 736 | 3-methyl-1-butanola | 0 | 0 | 0.1 ± 0.1 | 0–0.1 | 1.2 ±1.7 | 0.1–3.1 | 0 | 0 |
| 1267 | 1006 | 3,6-dimethyl-1,7-octadienea | 0.1 ± 0.1 | 0–0.2 | 0 | 0 | 0.1 ± 0.1 | 0–0.2 | 0 | 0 |
| 1445 | 1091 | Unknown3b | 0.2 ± 0.2 | 0–0.4 | tr | 0–0.1 | 2.4 ±4.2 | 0–7.2 | 0 | 0 |
| 1456 | 1380 | Unknown4b | 1.8 ±2.9 | 0–7.6 | 0 | 0 | 0 | 0 | 0.4 ± 0.7 | 0–1.4 |
| 1517 | 1398 | 2,6,10-trimethyl-1,5,9-undecatrienea | 0.5 ± 0.5 | 0–1.1 | 0.1 ± 0.1 | 0–0.2 | 0 | 0 | 0 | 0 |
| 1604 | 1086 | Hotrienola | 0.3 ± 0.3 | 0–0.9 | tr | 0–0.1 | 0 | 0 | 0 | 0 |
| 1632 | 1420 | Unknown5 | 0.3 ± 0.3 | 0–0.7 | tr | 0–0.1 | 0 | 0 | 0 | 0 |
| 1637 | 1451 | Alloaromadendrene | 0.5 ± 0.6 | 0–1.5 | 0.1 ± 0.1 | 0–0.1 | 0 | 0 | 0 | 0 |
| 1655 | 1180 | 3,6-dimethyl-1-octanola | 0.1 ± 0.2 | 0–0.4 | tr | 0–0.1 | 0.1 ± 0.1 | 0–0.2 | 0 | 0 |
| 1666 | 863 | 3-methylbutanoic acida | 0.2 ± 0.2 | 0–0.4 | 3.7 ±5.1 | 0.1–7.3 | 3.3 ±1.6 | 2.3–5.2 | 0.2 ± 0.2 | 0–0.5 |
| 1701 | 1472 | Germacrene D | 1 ± 1.4 | 0–3.6 | 0 | 0 | 0 | 0 | 0 | 0 |
| 2079 | 1052 | p-cresol | 7.6 ±4.1 | 3.3–14.1 | 22.9 ±17.6 | 6.2–39.6 | 17 ±12.3 | 5.4–29.9 | 5.6 ±3.2 | 3.2–10.3 |
| 2439 | 1252 | Indole | 3 ±2.9 | 0.6–10.1 | 1.4 ±1.8 | 0.1–2.7 | 1 ± 1 | 0.1–1.9 | 0.2 ± 0.3 | 0–0.5 |
| 2485 | 1344 | Skatole | 3 ±2.4 | 0–6.8 | 3.4 ±3.7 | 0.8–6 | 1.2 ±1.5 | 0.1–3 | 0.7 ± 0.2 | 0.6–1.1 |
Volatile compounds are classified by increasing order of their Kovats retention index (RI) on the polar column (BP-20), RI on the non-polar column (BP-1) are also given. Those with an average relative proportion above 1% are indicated in bold, and tr those with an average relative proportion below 0.1%.
aTentative compound identification.
bMass spectra given in the Supplementary Material.
Contrary to the previous published floral scent of A. pictum, we found noticeable differences. Among the five major VOCs found previously in Corsican inflorescences [7], two have also been found in the present study, namely indole and skatole. The three others were not identified in any of our samples from Corsica or Sardinia [7], (Table 2). The isomyocorene was probably mistakenly identified as ocimene, because the two compounds have similar framework and differ by a double bond, the first is 2,6-dimethyl-1,3,7-octatriene whereas the second is 3,7-dimethyl-1,3,6-octatriene. Also, p-cresol was probably mistakenly identified as benzyl alcohol, and β-citronellene, synonym of dihydromyrcene, as dihydroocimene. No Kovats retention indexes were published and several tentative compound identifications were performed by manual mass spectra interpretations [7].
Arum pictum is characterised by a dung odour close to horse/donkey dung [1]. Several foul-smelling VOCs were identified from the floral odour of A. pictum, most related to dung (as p-cresol, skatole, or indole) or fermentation/cheese (3-methyl-1-butanol, 3-methylbutanoic acid) in combination with floral VOCs (β-citronellene, 3,6-dimethyl-1-octanol, germacrene D). Interestingly several major VOCs found in the floral scent of A. pictum were identified as important VOCs emitted by horse dung such as p-cresol, β-citronellene, indole or skatole [14]. On the other hand, the major VOC, 4-methyl-3-heptanone, from horse dung was not identified in the floral blend of A. pictum [14]. Such diverse VOC combination is probably linked to the deceptive pollination strategy of Arum in order to attract insects associated to dung and/or fermenting matters [8, 9, 11, 12, 13].
3.4. Visiting entomofauna
The visiting entomofauna was dominated in all populations by Diptera, representing 69–78% when considering all the arthropods but 80–89% when considering only insects (Table 3). Secondary insect visitors were Coleoptera, particularly Staphylinidae (rove beetles, 4–16%). The average number of insects trapped per inflorescence varied among populations, with fewer in Baratz (3.9) than in Coti (6.4) and Diga (6.6). Interestingly, the floral visitors in the populations of Coti and Baratz were both highly dominated by Brachycera flies (respectively 75 and 80%) with Staphylinidae (respectively 10 and 16%) being secondary visitors (Table 3). On the other hand, while the floral visitors in the population of Diga were mainly Diptera, Nematocera were attracted in greater proportions (68%) and Brachycera were less frequent (25%).
Diversity of inflorescence-visiting arthropods for three populations of Arum pictum
| Population | Coti | Baratz | Diga |
|---|---|---|---|
| Region | Corsica | West Sardinia | East Sardinia |
| Insect Orders\Sample size | N = 26 | N = 30 | N = 8 |
| Brachycera (Diptera) | 126 | 93 | 13 |
| Nematocera (Diptera) | 10 | 3 | 36 |
| Coleoptera | 12 | 1 | 0 |
| Staphylinidae (Coleoptera) | 17 | 19 | 2 |
| Braconidae (Hymenoptera) | 2 | 0 | 2 |
| Othersa | 51 | 21 | 10 |
Total arthropod abundances per order or family.
aOthers represent the sum of unlikely Arum pollinators: Collembola (N = 8), Acari (N = 34), Salticids (N = 29), Blattodea (N = 1), Hemiptera (N = 1), Formicidae (N = 4), Isopoda (N = 1), Lepidoptera (N = 1), Psocoptera (N = 1), Thysanoptera (N = 2).
In a previous study, the main pollinators of Arum pictum were sphaerocerid flies (Brachycera) and staphylinid beetles [7]. Although we did not identify the Diptera to family level, many of the Dipteran Brachycera were sphaerocerids, confirming the same kind of visiting insect community in A. pictum inflorescences from north-west Sardinia (Baratz) and south-west Corsica (Coti and Sagone in [7]). On the other hand, the population of north-east Sardinia (Diga) presented a different visiting insect community dominated by Nematocera (Table 3). Psychodid and Chironomid flies (Nematocera) are known to be the main pollinators of other European deceptive Arum species [8, 9, 10, 11, 12, 13]. Interestingly, the floral scent in the population of Diga was the one with the lowest proportion of dung associated VOCs (∼6.5% in average) compared to the three other studied populations (14–28% in average; Table 2).
4. Conclusions
The autumn flowering Arum pictum has developed a deceptive pollination strategy combining a dark brown-purple spathe, floral heat production and scent emission mimicking probably horse/donkey dung, and attracting a coprophilous/saprophilous insect community composed mainly of diverse Diptera and staphylinid Coleoptera. Several foul-smelling VOCs related to dung (p-cresol, skatole, or indole) were reported in notable quantities. But surprisingly the main floral VOC was 2,6-dimethylocta-1,3,7-triene, also known as isomyocorene. Its odour is not known but probably not related to dung scent as other documented dimethyl-octatrienes (ocimenes) or acyclic hydrocarbon monoterpenes. Further studies are necessary to better understand the relationship between the floral scent composition in Arum pictum and the diversity of the attracted insects as it has been done recently for A. maculatum [11, 12, 13].
Declaration of interests
The authors do not work for, advise, own shares in, or receive funds from any organization that could benefit from this article, and have declared no affiliations other than their research organizations.
Acknowledgements
AA thanks her parents for supporting her and AG during this work, particularly by hosting them in Calangianus and accompanying them to the east-Sardinian studied site.