

Version française abrégée
Le rapport isotopique de l'oxygène des groupes carbonate et phosphate de l'émail dentaire (OTE) des mammifères reflète le O de l'eau du corps (OBW) [7,21], qui dépend du bilan des flux d'oxygène entrants (alimentation, respiration, consommation d'eau) et sortants (urine, fèces, expiration, évaporation) [6,26]. Pour les grands mammifères, l'eau météorique, sensible au bilan précipitation/évaporation [23], est la principale source d'oxygène et détermine alors la valeur du OBW [6]. En raison du caractère homéotherme des mammifères [13], il est possible de reconstituer les paramètres climatiques à partir de la composition isotopique de l'oxygène des tissus minéralisés, comme l'émail dentaire, sans correction de variations de température [6,23]. L'hydroxyapatite de l'émail, caractérisée par trois sources d'oxygène (carbonate, phosphate et hydroxyle) [7], résiste généralement mieux à la diagenèse que la dentine ou l'os, en raison d'une plus forte cristallinité [21,24]. Aux moyennes et hautes latitudes, le OMW est corrélé à la température de surface ; des relations empiriques entre les changements à long terme du OMW et de la température de l'air de surface en un lieu donné ont été établies [16,22]. La latitude, l'altitude, l'effet de continentalité, les sources d'humidité, la saisonnalité et le montant des précipitations peuvent modifier la signature initiale du OMW [22]. Quantifier ces effets permet de corriger les valeurs du OTE [22]. Sur une plus grande échelle temporelle, la température et le montant des précipitations sont les deux principaux paramètres modulant le OMW.
Nous avons étudié les variations de la composition isotopique de l'oxygène des carbonates (OC) de six espèces de rhinocérotidés de quatre sites du bassin d'Aquitaine couvrant la période 23–12 Ma : Grenade-sur-Garonne (MN1 ; , , altitude : 110 m), Montréal-du-Gers (Béon 1, MN4 ; , , altitude : 144 m), Sansan (MN6 ; , , altitude : 237 m), et Simorre–Villefranche-d'Astarac (MN7 ; , , altitude : 200 m) (Fig. 1). Nous avons effectué des analyses isotopiques sur vingt dents de rhinocéros (13 molaires et sept prémolaires : on utilisera respectivement M et P pour les dents jugales supérieures et m et p pour les inférieures) appartenant à six espèces rapportées à plusieurs sous-familles (Fig. 2). Du site de Montréal-du-Gers, des spécimens appartenant à quatre espèces de rhinocérotidés, dont une mandibule incomplète (deux prémolaires et deux molaires) d'un Prosantorhinus douvillei adulte, ont été intégrés à l'étude [1,14]. En termes de biochronologie mammalienne, les localités étudiées correspondent aux unités MN1, MN4, MN6 et MN7 (respectivement ∼23–22, ∼18–17, ∼15–12 et ∼12–11 Ma [35]). Sansan représente la localité de référence de MN6, avec un âge magnétostratigraphique compris entre 15,2 et 15,0 Ma [33]. Pendant le Miocène inférieur et moyen, la partie occidentale du Bassin aquitain a été envahie par la mer à plusieurs reprises [19,28], et tous les sites d'étude étaient, à l'époque, côtiers.
Location map of the four Miocene sites.
Carte des quatre sites étudiés du Miocène du bassin d'Aquitaine (SW France).
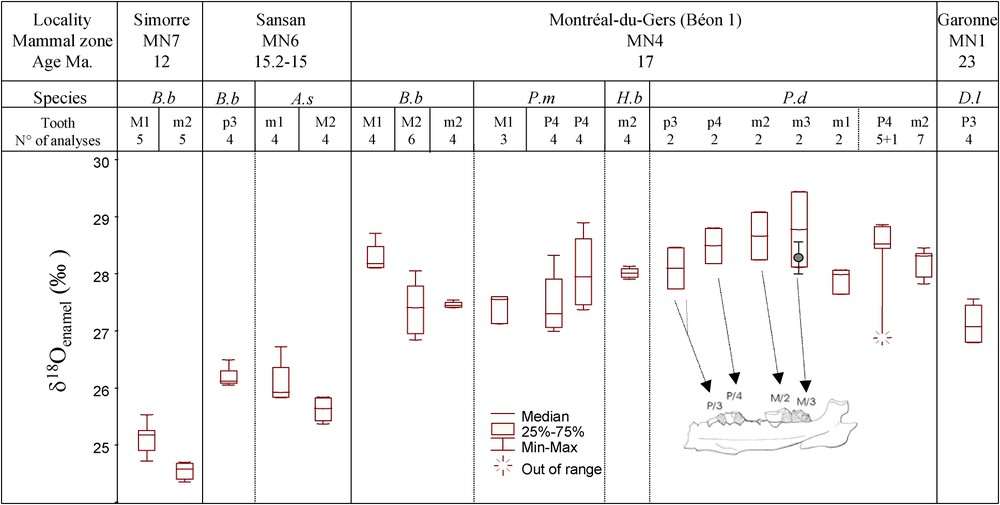
Box plots of the 20 mean teeth OC (V-SMOW) versus the sites and MN biozones, species (D.l, B.b, P.d, P.m, A.s, H.b refer to Diaceratherium lemanense, Brachypotherium brachypus, Prosantorhinus douvillei, Plesiaceratherium mirallesi, Alicornops simorrense, Hispanotherium beonense, respectively), tooth identification (abbreviations, l = left, r = right, P = upper premolar, p = lower premolar, M = upper molar, m = lower molar, referring to position in the tooth row), and number of analyses per tooth. The boxes, horizontal and vertical bars represent the 50% of the variability, the median and the amplitude of the intra-tooth OC, respectively. Inset in the Montréal-du-Gers section: the left hemi-mandible and associated teeth of a P. douvillei specimen. The oxygen isotopes of the phosphate group of the m3 of P. douvillei (italics) were also measured to check the enamel preservation. We converted the measured (N=4) into Oc-theoretical (grey dot) for comparison with measured Oc-measured (box plot).
OC des dents de rhinocéros en fonction des localités et biozones MN, des espèces (D.l, B.b, P.d, P.m, A.s, H.b se réfèrent respectivement à Diaceratherium lemanense, Brachypotherium brachypus, Prosantorhinus douvillei, Plesiaceratherium mirallesi, Alicornops simorrense, Hispanotherium beonense), des dents identifiées (chaque dent a été sous-échantillonnée en deux à sept fragments) et du nombre d'analyses par dent. Les boîtes représentent 50% de la variabilité, les barres horizontales à l'intérieur des boîtes représentent la médiane et les barres verticales représentent les valeurs extrêmes du OC d'une dent. Dans la section Montréal-du-Gers, une demi-mandibule d'un spécimen de P. douvillei, avec, en place, plusieurs dents, est représentée. Nous avons mesuré les isotopes de l'oxygène des phosphates de l'émail (Op) de la m3 de P. douvillei (en italique) comme un test de la diagenèse. Nous avons converti ces valeurs en Oc-théorique pour comparaison avec le Oc-mesuré.
Les valeurs de OC sont reportées sur la Fig. 2. L'intégrité cristallographique de l'émail a été contrôlée par l'analyse des spectres de diffraction des rayons X [30]. L'indice de cristallinité (IC) et la largeur à mi-hauteur des pics de diffraction montrent que l'ensemble des échantillons est bien préservé, à l'exception de la P4 de P. douvillei (Montréal-du-Gers) et de la P3 de D. lemanense (Grenade). La fraction carbonatée de l'émail est la plus sensible aux échanges isotopiques [34,37], ce qui se traduit par la perte de l'équilibre isotopique existant dans l'émail de mammifères entre le OP et OC. Un émail préservé est caractérisé par un fractionnement isotopique apparent entre l'oxygène des groupes phosphate et carbonate de l'émail de [7,21]. Une valeur de 8,97‰ sur la m3 de P. douvillei montre que la diagenèse n'est pas significative (Fig. 2).
Entre 18 et 12 Ma, la déplétion isotopique observée dans le bassin d'Aquitaine, situé à la paléolatitude 40°N [32], peut être liée à une baisse de la température de l'air si le montant des précipitations est resté stable [36]. Selon la relation de Dansgaard (pente de [16]), les variations isotopiques correspondent à une baisse de température variant entre 4,8 et 6 °C, en fonction des méthodes de calcul de OMW utilisées [5,21]. Ce refroidissement est en phase avec la baisse de pCO2 (au moins 100 ppmv [31]) entre 18 et 12 Ma. Utescher et al. [36] ont calculé une augmentation de 2 °C entre 23 et 17 Ma et une diminution globale de température de 3–4 °C entre 17 et 12 Ma. Miller et Sugarman [29] citent, pour le domaine marin, une augmentation des isotopes de l'oxygène de 1‰ entre 17 et 12 Ma, et trouvent une corrélation avec la baisse de 4–5 °C des températures des eaux de surface de la mer du Nord [9]. Nos données de OMW sont cohérentes avec la tendance globale des études paléocéanographiques [2] (Fig. 3a et c) concernant (i) la déplétion isotopique du Miocène inférieur à moyen, (ii) le début de l'expansion de la calotte antarctique entre 16 et 13 Ma et (iii) le changement dans la circulation d'eaux profondes entre 13 et 11 Ma, mis en évidence dans les séquences marines. Cependant, comme le montre la Fig. 3d, les températures d'eaux profondes, reconstituées à partir du rapport Mg/Ca des foraminifères benthiques, mettent en évidence (i) une variabilité climatique tout au long du Miocène moyen, marquée par des changements de température de l'ordre du degré et (ii) au moins deux réchauffements importants culminant à 12 et 8,5 Ma, ce qui ne concorde pas avec notre interprétation initiale d'un refroidissement progressif (toutefois, nos données sont éparses) entre 18 et 12 Ma. La bonne correspondance de nos données avec le O de l'eau de mer [2] (Fig. 3c) accompagnant ces événements chauds suggère, par déconvolution du signal, que les précipitations sont un facteur forçant. Cette étude a le mérite, dans le contexte scientifique actuel, de remettre au premier plan l'importance du rapport évaporation/précipitation sur le signal O (en domaine continental aussi bien qu'en domaine marin). En perspective, il faut (i) croiser nos données avec d'autres données continentales indépendantes (par exemple, la faune, le pollen), (ii) améliorer significativement la résolution temporelle des données continentales et (iii) appréhender quantitativement les changements saisonniers potentiels des paramètres climatiques [17].
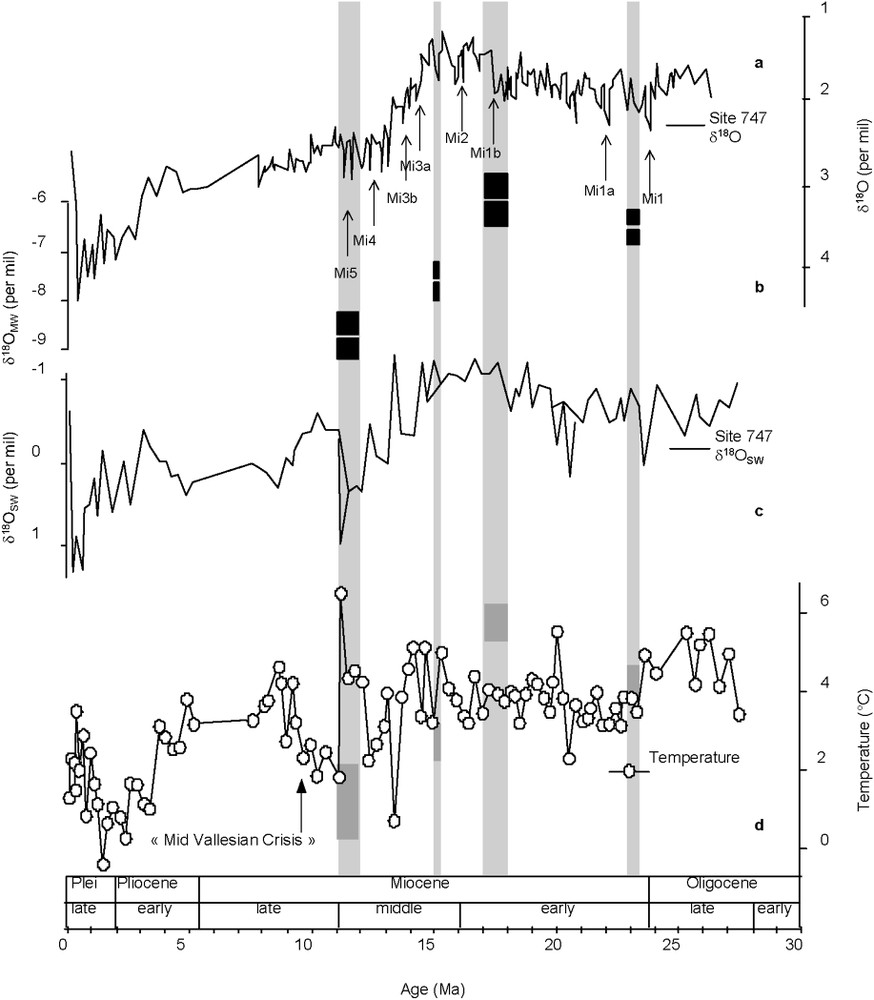
Following Billups et Schrag [2]: (a) O of the benthic foraminifera Cibicidoides sp. [39]; (b) average OMW (black rectangles); (c) OSW of seawater [2] and (d) site 747 deep-water reconstructed temperature [2]; the grey rectangles correspond to the reconstructed palaeotemperatures (expressed in Δ) obtained from the Aquitaine Miocene OC.
Modifiée d'après Billups et Schrag [2] : (a) O de foraminifères benthiques Cibicidoides sp. [39] ; (b) valeur moyenne du OMW (rectangles noirs) ; (c) OSW de l'eau de mer [2] et (d) température des eaux profondes, reconstruite au site 747 [2] ; les rectangles grisés correspondent aux reconstructions des paléotempératures (exprimées en Δ, anomalies par rapport à l'Actuel) obtenues à partir des OC des sites Miocène d'Aquitaine.
1 Introduction
Carbonate and phosphate oxygen isotope ratios of mammal tooth enamel (OTE) provide information on the oxygen isotope ratio of body water (OBW) [7,21] in turn related to the oxygen fluxes that enter (food, respiration, drinking) and leave the body (urine, faeces, expiration, evaporation) [6,26]. For large-sized mammals such as rhinocerotids, the main oxygen source determining OBW is meteoric drinking water (OMW) [6], sensitive to the precipitation/evaporation balance [23]. Because mammal tooth enamel is formed at a constant body temperature [13], there is no need for temperature corrections [6,23]. Therefore, the OTE is mainly correlated to the OMW [15,23,26]. Since enamel hydroxyapatite (Ca10(PO4, CO3)6(OH, CO3)2 [7], characterized by three oxygen sources) is generally more resistant to diagenetic alteration than dentine and bone [21,24], the OTE is an adequate proxy of past OMW [5,6,8,34]. In mid and high latitudes, OMW in monthly and long-term precipitation changes are correlated with the surface-air temperature [16,22]. Latitude, altitude, continental effect, humidity sources, seasonality and the amount of precipitation also influence the OMW [22]. Quantification of some of these effects corrects OTE for oxygen source variability [22]. Over a long-term period, both temperature and precipitation rate are the main parameters modulating OMW.
In this study, oxygen isotopic compositions of enamel carbonate (OC) from six rhinocerotid species and four mammal localities (Early to early Late Middle Miocene, 23–12 Ma) are measured. Montréal-du-Gers contained four species of coexisting rhinocerotids [1,14]. We analysed the intra-, inter-tooth and inter-species (contemporary fossils) OC variability, and the OC time dependence. Results are compared to palaeontological and palaeoceanographic data.
2 Sampling and analytical techniques
Four Miocene mammal localities in Aquitaine (France) were studied (Fig. 1): Grenade-sur-Garonne (MN1; , altitude: 110 m), Montréal-du-Gers (MN4; , altitude: 144 m), Sansan (MN6; , altitude: 237 m), and the Simorre–Villefranche-d'Astarac ensemble in the ‘Astarac’ area (MN7; , altitude: 200 m). Twenty rhinocerotid teeth (13 molars; 7 premolars) were studied. Six species were identified: (i) three teleoceratines: middle-sized Diaceratherium lemanense (Pomel, 1853), large Brachypotherium brachypus (Lartet, 1837), and small-sized Prosantorhinus douvillei (Osborn, 1900): hippo-like rhinos adapted to swamps and riversides [10] with short limb bones and brachyodont cheek teeth; (ii) the aceratheriines Plesiaceratherium mirallesi (Crusafont, Villalta and Truyls, 1955) and Alicornops simorrense (Lartet, 1851). They were more slender-limbed, hornless rhinos adapted to long distance running, probably in open woodlands (brachyodont teeth); (iii) elasmotheriine Hispanotherium (Aegyrcitherium) beonense [1], characterised by slender limb bones and high crowned teeth filled with cement. This species is thought to be a mixed browser-grazer living in open environments. D. lemanense (Earliest Miocene), P. mirallesi, H. beonense and P. douvillei (late Early Miocene) have short stratigraphic ranges and are restricted to western Europe. A. simorrense (Middle to Late Miocene) and B. brachypus (late Early to Late Miocene) occurred widely in Europe and Anatolia [10]. In the European Neogene mammal chronology our fossil content corresponds to units MN1 (∼23–22 Ma), MN4 (∼18–17 Ma), MN6 (∼15–12 Ma) and MN7 (∼12–11 Ma), respectively [35]. Sansan is the reference locality for unit MN6, with a magnetostratigraphic age between 15.2 and 15.0 Ma [33].
West European palaeoenvironments were affected by two major tectonic events between 17–14 and 9–8 Ma [28]. During the Early and Middle Miocene, the western part of the Aquitaine region has been invaded by the sea several times [19], and the four localities were close to their contemporary coastline. The ‘Astarac region’ (MN7) faunas are characterized by the co-occurrence of B. brachypus and A. simorrense. The environment was most likely a wet forest. At Montréal-du-Gers (MN4), fossils were collected from clay beds representing a wet paludal and/or lake-side environment, surrounded by a wooded savannah-like biome where H. beonense, P. mirallesi, B. brachypus, and P. douvillei occurred [38]. In this locality, a jaw with an incomplete tooth row (two premolars and two molars) belonging to an adult P. douvillei was studied. A single specimen of D. lemanense found in Grenade-sur-Garonne (MN1) was sampled.
Between 2 and 7 fragments were analysed for each tooth, giving an indication of intra-tooth variability.
Diagenesis tests were used in order to check both the crystallinity and isotopic integrity of the fossil teeth. Scanning Electron Microscopy observations showed that crystalline structures are preserved. The constant crystal index for the fossil teeth and full width half maximum as measured by X-ray diffraction (XRD) analysis [30] show that all of the fossil teeth (except the P4 [P. douvillei, MN4] and the P3 [D. lemanense, MN1]) are well preserved. PO chemical bonds in apatite are more resistant to alteration process than the CO bonds [34,37], so phosphate oxygen isotopic composition (OP) is often preferred to that of carbonate [24]. However, the OP and OC isotopic differences can be used as a benchmark of the fossil preservation state. Diagenetically altered specimens may show deviations of OC from the expected line [21]. We have tested a single tooth (m3, P. douvillei from Montréal-du-Gers); the difference between OC and OP is 8.97‰ (corresponding to an apparent carbonate–phosphate fractionation factor of , in good agreement with previous work [7,21]), which supports the conclusion of no significant diagenetic alteration.
The teeth were drilled either on appropriate parts of broken surfaces of the same tooth or along the labial or lingual surface of a loph(id) perpendicular to the occlusal plane. Eighty isotopic analyses were performed. Five milligrams of powdered enamel were obtained using an agate mortar and pestle, sieved through a 125-μm mesh. Organic matter was removed using standard procedures [11]. Washed and treated enamel powder (1.5–2 mg) is acidified with phosphoric acid (103%) releasing CO2 gas for isotopic analyses using a Micromass Optima mass spectrometer. O is expressed against the international standard V-PDB with a precision better than . We converted the OC(PDB) into OC(V-SMOW) using the equation [12]: OV-PDB = 0.97001 × δ18OV-SMOW − 29.99.
3 Results and discussion
The results of the isotopic analysis are presented in Fig. 2, together with the provenance, species and number of analyses for each tooth. The box-plots exhibit a general decrease of the OC from the late Early to the Middle Miocene .
3.1 Intra-individual variability: P. douvillei
No significant trend (Fisher test: ) in the inter-tooth OC ranges of four teeth from a single jaw of P. douvillei found in the MN4 biozone has been observed (from p3 to m3, mean OC are and , respectively), in agreement with the m1 and m2 OC of other specimens of P. douvillei from the same deposit. The outlying P4 reflects either a diagenetic alteration, as suggested by the XRD, or a sampling artefact, such as dentine contamination. The lesser OC value of the m1 represents probably a physiological effect related to the influence of mother's milk [18]. All P. douvillei teeth OC (except P4), vary from 27.6 to 29.4‰. The 1.8‰ amplitude is probably a smoothed signal of the seasonal variability of the ingested oxygen (drinking water or another source). The sequence from the first to the last erupted tooth in mammals ranges from a few months to about two years [20]. Therefore, the OC of a tooth eruption sequence integrates the climate experienced by the growing mammal from weaning through to adulthood. However, the interannual variability of the isotopic signature (short-time scale) is likely to be far smaller than that of long-term geological climate change. Whatever the type of tooth for one single species found in a mammal locality, it should always give an accurate climatic signal.
3.2 Inter-species variability
The average OC and standard deviation of the four species from the MN4 biozone are , , and for B. brachypus, P. mirallesi, H. beonense and P. douvillei teeth, respectively. Three out of four thus have a similar average isotopic signal. The value of P. douvillei is heavier. The m2 mean OC of H. beonense is not significantly different with respect to the three other species, although the sample number is limited. P. mirallesi and B. brachypus teeth exhibit similar OC, significantly different compared to P. douvillei. The B. brachypus and A. simorrense OC from Sansan (MN6) are similar.
The species recovered from Montréal-du-Gers lived in different contemporary ecosystems: swamps (B. brachypus and P. douvillei), open woodland (P. mirallesi) and savannah-like open environments (H. beonense). The similarity of the OC suggests a common source of drinking water for all. B. brachypus (swamps) and A. simorrense (open woodlands) from the MN6 confirm this assumption. However, B. brachypus and P. douvillei (MN4) show that two rhinos thought to share the same habitat can have different OC. Three hypotheses are advanced: (i) the individuals were not strictly contemporary, but lived at intervals of several decades, (ii) the specific vital effect such as the evaporation rate (higher for P. douvillei than for B. brachypus suggesting that B. brachypus would have a typical hippo-like metabolism reducing evaporation), or (iii) an ingestion effect such as the oxygen content of preferred leaves or grasses [27].
3.3 Variability with respect to the geological age
The OC values vary between 24.4 and 29.4‰ for the period 23–12 Ma. The average OC value of the D. lemanense tooth in the MN1 biozone is , depleted by about 1‰ compared to the average value of the teeth of the four species from the MN4 biozone . There is evidence for crystallite diagenesis for this sample; however, crystal diagenesis and isotopic stability are not necessarily contradictory [25]. The 1‰ difference may be significant of climatic or interspecies change. The MN1 biozone corresponds to the end of the cooler Oligocene times and the beginning of the warmer Miocene times [36]. In MN6 and MN7, the OC becomes significantly more depleted, with an average of and , respectively.
In order to isolate the effect of the geological age on the variability of the OC, we have grouped the OC values from the same site (MN7: B. brachypus mean OC; MN6: B. brachypus and A. simorrense mean OC; MN4: B. brachypus, P. mirallesi, and H. beonense mean OC), and analysed B. brachypus from the MN4 through to the MN7 biozone. The single species B. brachypus OC results show significant differences. The averaged values are , and , respectively, at 18–17, 15.2–15.0 and 12 Ma. The 2.9‰ amplitude variation indicates palaeoenvironmental changes as a function of time. As there is no reliable calibration relating the rhino OC to the OMW, we have used Iacumin's carbonate–meteoric water calibration as the best approximation [21]. The OMW values for the period 17–12 Ma vary between and (Fig. 3b). As several calibrations between OMW and OP are available for large mammals [5], the OMW can also be estimated by transforming the OC values into OP and then deducing the OMW from the OP. Calculated average OMW decrease from to . The second method shows a larger isotopic shift of 3.6‰ between 17 and 12 Ma. Bryant and Froelich [5] found 5‰ for the period between 18 and 8.5 Ma in Nebraska (from −6.7 to ). The relationship between OMW and temperature derived for modern precipitation [16] is not strictly applicable to palaeoclimate reconstruction. However, it is possible to reconstruct ΔT (°C) (anomalies with respect to the present). These isotopic variations could therefore be due to a cooling and/or an increase in precipitation.
3.4 Isotopic shift and temperature changes
If we assume that the observed isotopic shift is mainly a temperature effect at this palaeolatitude of about 40°N [32] and that the precipitation amount remained stable throughout the period of our study, as shown for the Neogene of western Europe [36], then we estimate – using Dansgaard's calibration of [16] – a 4.8 to 6 °C cooling between 18 and 12 Ma, in phase with a pCO2 drop (100 ppmv [31]). Utescher et al. [36] calculated a 2 °C increase from the Late Oligocene–Early Miocene (23–17 Ma) and a global temperature decrease between 17 and 12 Ma of about 3–4 °C. Miller and Sugarman report a 1‰ marine oxygen isotope increase between 17 and 12 Ma [29] correlating it to a shift of about 4–5 °C of the North Sea sea-surface temperature [9]. Our data concur with the global pattern given by palaeo-oceanographic studies [2] (Fig. 3a–c) concerning (i) the Early to Middle Miocene isotopic depletion, (ii) the onset of the East Antarctic ice cap between 16 and 13 Ma, and (iii) the deep sea sediment sequences showing deep water circulation changes between 13 and 11 Ma. However, Mg/Ca-based deep-water temperature reconstruction ([2], Fig. 3d) shows (i) a Mid-Miocene climate variability characterized by temperature changes of ca 1 °C, (ii) the occurrence of at least two major warming events culminating at ∼12 and 8.5 Ma, contradicting a progressive cooling as suggested by our sparse data (between 18 and 12 Ma) alone. Comparison with Billups and Schrag's O sea water and reconstructed temperature curves [2] thus suggests that precipitation changes had a major influence during the Mid-Miocene, as suggested by palaeontologists [3,4] at ca. 14 Ma.
In order to reconstruct more precisely Tertiary palaeoclimates, we will have (i) to cross our geochemical data with other independent continental proxies (fauna, pollen), (ii) to increase significantly the time resolution sampling of the continental records, and (iii) to reconstruct quantitatively the changes in terms of seasonal distributions [17].
Acknowledgements
We thank Mr R. Guiraux (teeth preparation and technical advice) and Mr D. Excoffon (XRD and SEM analyses), Dr. S. Papadimitriou and Dr D. Kroon for comments on a preliminary draft. This is the ISEM contribution No. 2005-093.