

1 Introduction
Testate amoebae are ubiquitous protists that live in various environments (peat land, rivers, lakes, brackish-water settings…) (Charman, 2001). The analysis of these organisms is now a common tool in paleoecological studies of lakes (Burbidge, 1998; Ellison, 1995; Medioli and Scott, 1983).
However, as all environmental parameters that govern species distribution are still imperfectly known, the interpretation of microfossil data remains equivocal. One bias may stem from the ecological conditions prevailing during test construction. Many factors control testate amoebae abundance and assemblage types, including nutrition, dissolved oxygen conditions, pH, salinity, substrate, temperature, subaerial exposure, or floral substrates (Scott et al., 2001). One largely unexplored factor is the sediment grain size and mineralogical composition. The texture of agglutinated tests may be highly variable. Agglutinated testate amoebae have the ability to select grains of different shape and size to make the three-dimensional form of the test. Some species are known to select the grains in the sediment (Eckert and McGee-Russell, 1974). The nature of the agglutinated tests may depend on their sedimentological environment, which may introduce some constraints and modifications in the structure of assemblages.
A preliminary study on the discrimination of mineral particles in test formation was carried out by Stout and Walker (Stout and Walker, 1976). X-ray qualitative analysis allowed the authors to precise that variations exist in the composition of the tests in the size range of the particles and their chemical composition. Nevertheless, very few mineral grain measurements were given and chemical analysis was about chemical-element detection and not mineral reconstruction. There is no clear validation whether testate amoebae depend of their substrate sediment to live and develop.
The aim of this study is to investigate the grain size and mineralogical assemblage of tests of common species belonging to the genus Centropyxis and Difflugia, commonly present in lakes, and to compare them with the sediment parameters of their substrate. This objective is a prerequisite before using testate amoebae for past reconstruction of climate and environment, for example.
2 Material and methods
2.1 Study area
Three lakes were selected in the Subalpine Massifs of the French Alps (Fig. 1): Annecy, Anterne and Gers lakes. Their surface areas are 26.5 km2, 0.12 km2 and 0.064 km2, respectively. The lakes collect water from watersheds of 251 km2, 5.5 km2 and 5.4 km2, respectively.
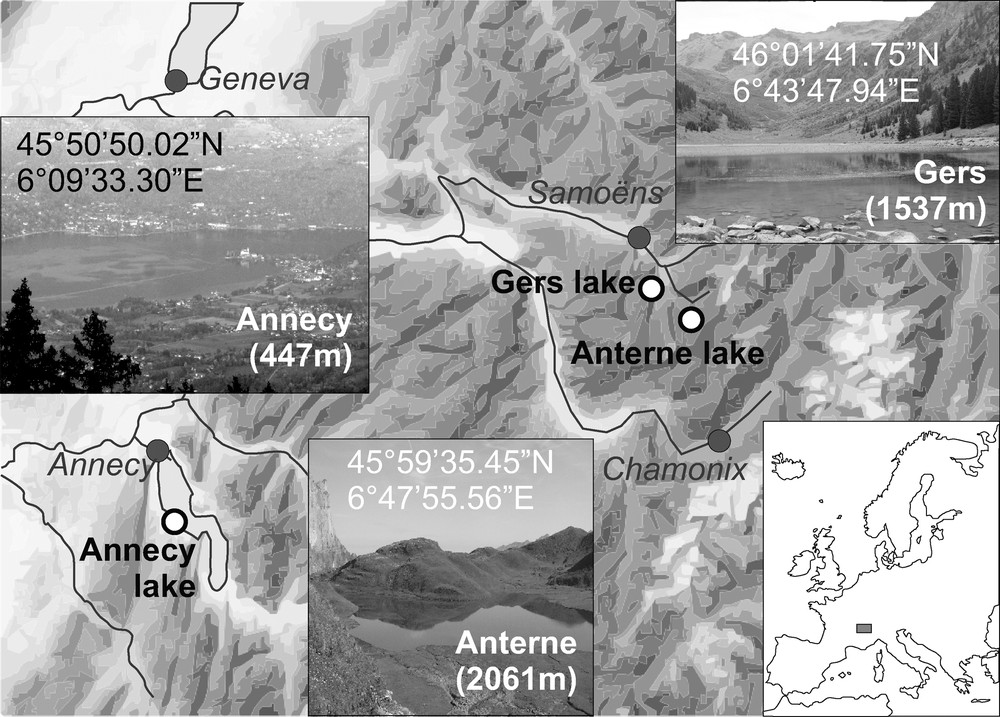
Study area and location of the sampling stations.
Zone d’étude et localisation des stations de prélèvement.
Annecy and Gers are oligo-mesotrophic and Anterne is oligotrophic (Druart and Balvay, 2009; Jochenbeim, 2002). The lakes are located at different altitudes, 447, 2061 and 1537 m, respectively. The rocks surrounding their watersheds drain surrounding rocks of contrasted lithologies, as reflected by the sediment composition. Alpine meadow is partially covering those rocks.
2.2 Sampling method and testate amoebae analysis
In this preliminary study, we collected in each lake one sample in August 2008 that was relatively voluminous with respect to the size and abundance of testate amoebae (further developments will require a spatial sampling). For each station, a constant volume of 20 cm3 of sediment was sampled from a 1/4 square meter zone, following a pseudo-replication method that allowed a bypass of the short scale patchiness (Debenay and Guillou, 2002). In the laboratory, wet samples were washed through 20 and 315 μm mesh sieves. Observations were carried out directly on the sediment of the intermediate fraction (20–315 μm) under a Olympus SZX16 binocular allowing a magnification of ×115.
Thirty-two specimens of agglutinated testate amoebae were selected because of their abundant occurrences in the three lakes (8 specimens at Annecy, 9 at Anterne and 15 at Gers). These specimens belong to nine species (Centropyxis aculeata, C. aculeata var. discoides, C. constricta, Difflugia corona, D. labiosa, D. oblonga, D. protaeiformis, D. pyriformis and D. urceolata).
The determination of the minerals agglutinated within the test walls of testate amoebae was performed by means of punctual analysis using Raman microspectrometry and an Environmental Scanning Electron Microscope (ESEM) equipped with an Energy Dispersive Spectroscopy (EDS) device. With ESEM-EDS, tests were imaged for elemental mapping (Si, Al, Fe, Mg, Ca, Na, K, Ti, and also S, C or Mn when detected) under low-vacuum conditions, using a 15 kV beam for point analyses of individual grains. Chemically coded color imaging helped separating grains according to their chemical composition, while conventional BSE-imaging helped localizing heavy element-rich grains (typically chlorite, rarely titanite or pyrite) because of their stronger reflectivity. Mineralogical composition was determined through chemical mapping and image analysis (mainly quartz vs. feldspar vs. sericitous mica).
The mineralogical composition of the tests was compared to similar analyses carried out on thin sections after resin impregnation of the sediment.
The grain size of the various minerals building the tests was directly measured on the ESEM-EDS maps. The substrate-sediment grain size was characterized using a Malvern Mastersizer 2000 (size range 0.02–2000 μm) that allowed determining modes, sorting and asymmetry.
3 Results
3.1 Chemical analysis
3.1.1 Mineral evaluation
Due to the small grain size compared to the volume analyzed by the EDS probe (about one cubic micrometer), an influence of the compositions of the adjacent grains is possible and cannot be distinguished from a possible contamination effect by the organic binding cement. As an example in Fig. 2, the measured Al-, Na- and K-contents (de la Roche, 1978) are combined and compared to the same parameters for ideal feldspar end-members and phengitic micas of the muscovite-celadonite series. All the parameters are nearer to 0 than pure minerals. This pattern can be related to the neighboring effect. In the case illustrated in Fig. 2 (Centropyxis constricta test from the Gers Lake), the analytical contamination might come from the binding cement and/or a thin carbonate layer coating the test.

ESEM-EDS grain analyses of one specimen Centropyxis constricta test from Gers Lake. Al/3-Na and Al/3-K as atomic ratios after de la Roche (1978), with the feldspars + quartz tetrahedron, using structural formulae for 11 oxygen (with Fe as Fe2+). Conventional mineral symbols are used (after Kretz (1983)), except for: Phg, phengite – Cel, celadonite.
Analyse des grains minéraux d’un individu de Centropyxis constricta du lac de Gers à l’aide d’une sonde EDS au MEB Environnemental. Al/3-Na et Al/3-K sont les rapports atomiques définis d’après de la Roche (1978), avec un tétraèdre feldspaths + quartz, utilisant des formules structurales pour 11 oxygènes (avec Fe pour Fe2+). Les symboles minéraux conventionnels sont utilisés (d’après Kretz (1983)) sauf pour: Phg, phengite - Cel, celadonite.
3.1.2 Sediment composition
In the Annecy Lake, the sediment studied is mainly composed of calcite whatever the size of the grains analyzed. Numerous large quartz and K-feldspar (microcline) minerals were observed. A few small grains of these minerals were also encountered. Very rare minerals of chlorite/Fe-oxide, albite, rutile, garnet and serpentine were observed.
In the Anterne Lake, the sediment is mainly composed of large quartz grains, accompanied by sericite, muscovite, calcite, Fe-oxide and rutile minerals. Two sericite types are distinguished, one with a composition close to muscovite (hereafter labeled Al-sericite) and a second with a composition close to celadonite, enriched in Fe, Mg and Si, and depleted in Al (labeled (Fe, Mg, Si)-sericite). All these minerals are also observed amongst the smallest grains.
The mineral composition of the sediment of the Gers Lake is similar to that observed from the Anterne Lake, with slightly more abundant mica.
3.1.3 Test mineralogical composition
Over the three lakes and the thirty-two specimens, the mineralogical nature of 5997 test-building grains was determined. Whatever the species, their overall composition is largely similar to that of the surrounding sediment (Table 1), namely, dominant calcite in the test of Annecy testate amoebae and a large proportion of quartz and mica in Anterne and Gers specimens. However, some inter- and intra-specific variability is observed (Table 1). For example, in the Anterne lake, where sediment is dominated by quartz, 96.4% quartz but no calcite is observed in Centropyxis aculeata whereas the test of Difflugia pyriformis is composed of 16.1% quartz and 63.7% calcite (inter-species variability). Within a same species, mineralogical composition is not constant and standard deviation of ratios can be very high (Table 1). For example, at the Anterne lake, Centropyxis constricta is composed of 14.2 ± 9.9% Fe-oxide, 43.1 ± 40.9% quartz, 27.4 ± 19.3% Al-sericite and 11.7 ± 10.5% (Fe, Mg, Si)-sericite.
Nombre de thécamœbiens analysés. Composition minéralogique des différentes espèces. L’écart type est donné quand c’est possible.
| Centropyxis aculeata | Centropyxis aculeata var. discoides | Centropyxis constricta | Difflugia corona | Difflugia labiosa | Difflugia oblonga | Difflugia protaeiformis | Difflugia pyriformis | Difflugia urceolata | |||
| Number of analyzed specimens | Annecy | 2 | 5 | 1 | |||||||
| Anterne | 1 | 2 | 3 | 1 | 1 | 1 | |||||
| Gers | 1 | 2 | 3 | 6 | 1 | 1 | 1 | ||||
| Mineral composition | |||||||||||
| Annecy | Chlorite/Fe oxyde | 3.4 ± 1.9 | 1.7 ± 1.7 | 0.0 | |||||||
| Quartz | 21.2 ± 18.8 | 7.5 ± 7.3 | 0.2 | ||||||||
| Microcline | 6.0 ± 3.5 | 7.0 ± 5.8 | 0.0 | ||||||||
| Albite | 1.9 ± 1.9 | 1.8 ± 3.4 | 0.0 | ||||||||
| Calcite | 67.5 ± 26.1 | 81.3 ± 10.0 | 99.8 | ||||||||
| Rutile | 0.0 ± 0.0 | 0.7 ± 1.6 | 0.0 | ||||||||
| Anterne | Fe-oxyde | 0.4 | 0.0 ± 0.0 | 14.2 ± 9.9 | 0.0 | 1.4 | 2.3 | ||||
| Quartz | 96.4 | 13.7 ± 0.2 | 43.1 ± 40.9 | 42.0 | 51.5 | 16.1 | |||||
| Sericite-(Al) | 3.2 | 8.9 ± 3.2 | 27.4 ± 19.3 | 6.7 | 3.6 | 6.0 | |||||
| Sericite-(Fe, Mg, Si) | 0.0 | 77.1 ± 3.5 | 11.7 ± 10.5 | 51.3 | 34.7 | 63.7 | |||||
| Calcite | 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 | 8.8 | 2.0 | |||||
| Rutile | 0.0 | 0.3 ± 0.4 | 0.9 ± 1.5 | 0.0 | 0.0 | 0.0 | |||||
| Muscovite | 0.0 | 0.0 ± 0.0 | 2.7 ± 4.8 | 0.0 | 0.0 | 9.8 | |||||
| Gers | Fe-oxyde | 1.0 | 0.0 ± 0.0 | 14.8 ± 3.3 | 9.1 ± 20.5 | 10.3 | 0.7 | 1.6 | |||
| Quartz | 21.6 | 15.0 ± 0.9 | 10.7 ± 5.0 | 29.6 ± 10.6 | 33.0 | 37.2 | 23.8 | ||||
| Sericite-(Al) | 8.2 | 7.7 ± 0.6 | 17.2 ± 9.4 | 25.1 ± 30.9 | 11.0 | 54.2 | 8.2 | ||||
| Sericite-(Fe, Mg, Si) | 69.2 | 50.1 ± 0.6 | 47.0 ± 12.5 | 36.1 ± 33.5 | 45.5 | 7.9 | 50.0 | ||||
| Calcite | 0.0 | 27.2 ± 1.0 | 9.6 ± 10.2 | 0.0 ± 0.0 | 0.3 | 0.0 | 16.4 | ||||
| Rutile | 0.0 | 0.0 ± 0.0 | 0.8 ± 1.3 | 0.0 ± 0.0 | 0.0 | 0.0 | 0.0 |
3.2 Grain-size measurements
The sediment from the three lakes have a similar unimodal grain-size distribution. The mode corresponds to fine sand at Annecy (56.7 μm) and coarse silt at Anterne and Gers (88.8 and 82.9 μm, respectively) (Fig. 3). Grains are poorly sorted (sorting between 1.5 and 2) and distributed with an asymmetry towards larger size. Small grains are scarce.
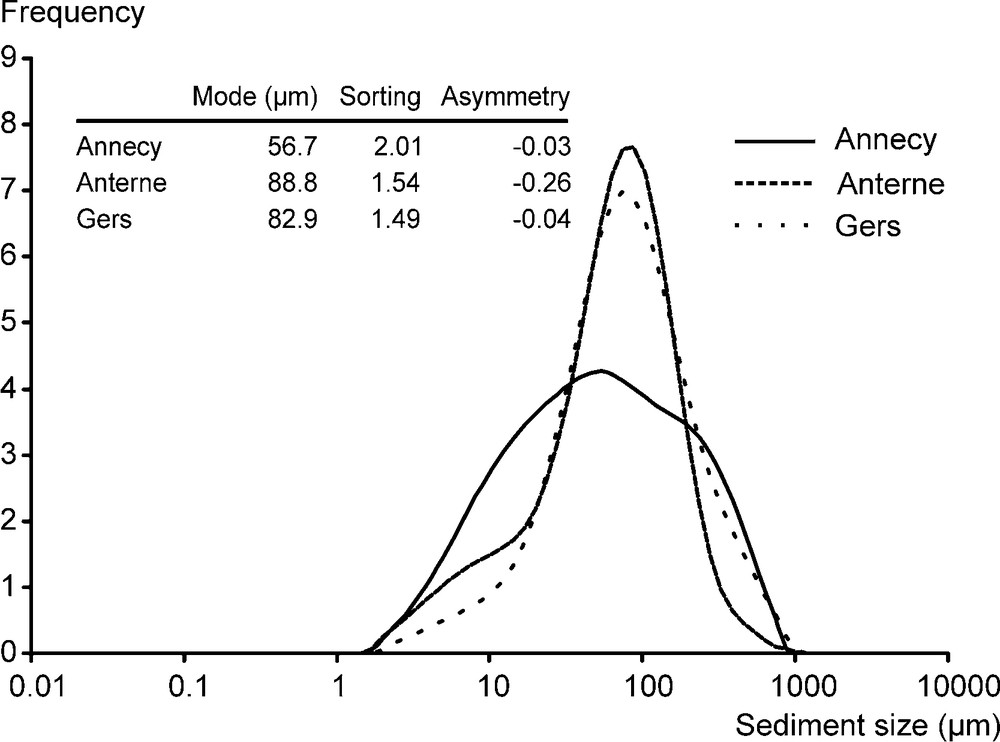
Grain size of the sediment of the 3 lakes.
Taille granulométrique des sédiments des 3 lacs.
The size of the grains agglutinated to form the test of the testate amoebae is largely small, with a median size smaller than 5 μm (Fig. 4a–c). The grain size of the Annecy specimens is smaller than that of the Anterne and Gers specimens, although the Annecy sediment is coarser than that of the other lakes. The grain size of the tests shows some intra-specific variability. For example, within Centropyxis constricta observed in the three lakes, quartz size is centered at ca. 1.5 μm at Annecy, ca. 3.5 μm at Anterne and ca. 2.5 μm at Gers (Fig 4a–c). At Annecy, amongst the dominant calcite grains in the sediment, those of 1.5 up to 2.5 μm seem to be specifically selected by the protists (Fig. 4a). The first and third quartiles are close to 1 μm in the analyzed three species. When grains are less abundant, as is the case with albite or microcline, the sizes of the grains selected by the protists are more variable. At Anterne and Gers, the mineral composition of the smallest grains available for test construction is more diverse than at Annecy.
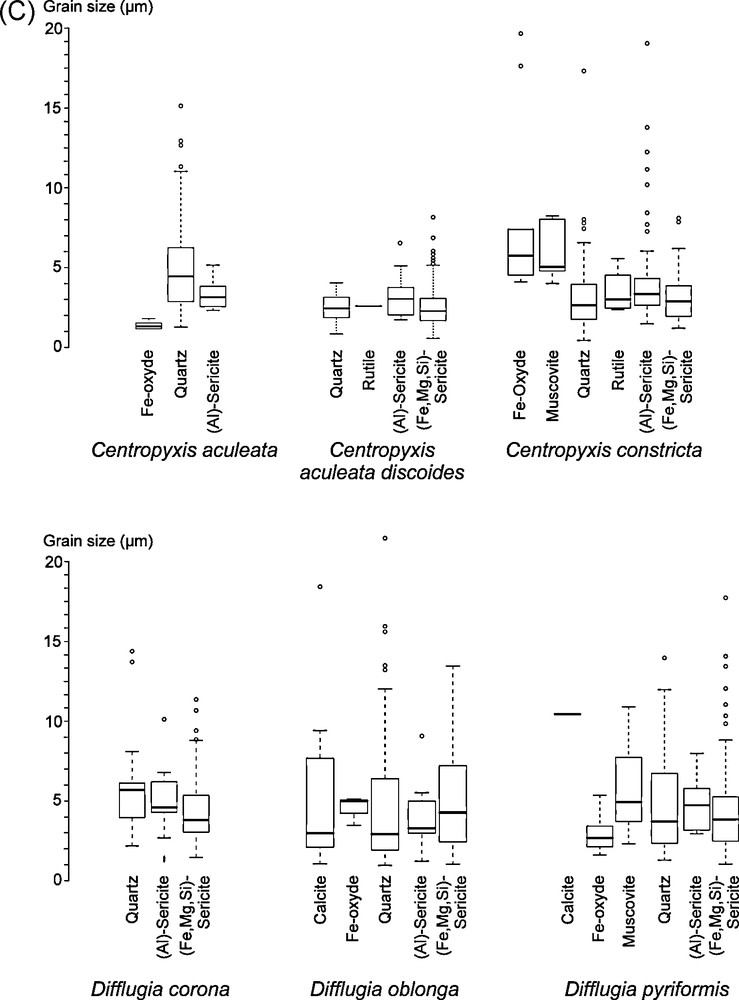
Size of the grains of the different minerals of each species of the 3 lakes: a = Annecy, b = Anterne and c = Gers.
Taille des grains des différents minéraux de chaque espèce des 3 lacs: a = Annecy, b = Anterne and c = Gers.
The size of grains is not homogenous within a same specimen. The aperture is often surrounded with grains that are smaller and better-sorted that those of the rest of the test (see ESEM picture of Difflugia oblonga in Fig. 5).

Log normal grain distribution from external areas of Centropyxis constricta and Difflugia oblonga.
Distribution log-normale des grains des surfaces externes de Centropyxis constricta and Difflugia oblonga.
When plotting Log (Frequency of grains) vs. Log (Mean grain radius), a Log-normal grain distribution is observed in all specimens. The mean mode is located at ca. 1.47 ± 0.66 μm. When small grains are scarce, the “pavement” is loose, implying that the fraction of organic cement binding the grains together is high (Fig. 5, Centropyxis constricta). Conversely when small grains are not a limiting factor, they are employed to fill the gaps between the other grains (Fig. 5, Difflugia oblonga).
4 Discussion
Our observations underline the dominating influence of the mineralogical composition of the sediment on the testate amoebae tests: the same species are present in sediment with contrasting mineralogy and their tests accordingly show contrasting mineralogical compositions. This observation suggests that sediment mineralogy is not a limiting factor for test construction. These protists can adapt to various substrate compositions. In the same way, it is known that cavities within organic matrix may harbor mineral grains that provide a more robust construction in Centropyxis discoides (Ogden, 1988a) or C. kahli (Ogden, 1988b). Testate amoebae may develop even where mineral grains are not present (Ogden, 1988b). In that case, their test is organic-walled.
However, some variability, either inter- or intra-specific, is observed relatively to the mean sediment composition, which must be accounted for. The mean grain-size distribution of the sediments studied here is different from that of the observed tests. Eckert and McGee-Russell (Eckert and McGee-Russell, 1974), concluded that the pattern of particles in shells of Difflugia might be specific. Our result is not supporting this point. The protists appear to be selective with regards to the size of the grains they retain to build their test. In addition, small-sorted grains are locally observed around the aperture of Difflugia corona, such as already observed around the apertural zone of Centropyxis arenaria (Golemanski and Ogden, 1980).
Furthermore, the test particles are smaller than the mean grain size of the sediment. Inter- and intra-specific variability is weak as far as test grain size is concerned with regards to grain mineralogy. In other words, our results strongly suggest that testate amoebae are rather selective with the size of the particles they use, but much less so with the mineralogical nature of these particles. The fact that they choose small-dimension grains within the entire size range of available grains, explains why the tests may have a different mineralogy and a mean grain size with respect to the substrate. As the prey selectivity is carried out either by physical contact or chemotactism (e.g., Gilbert (Gilbert et al., 2000)), it could be argued that these mechanisms may also occur in the test construction. The grain-size selectivity coupled to some indifference toward mineralogy indicates that chemotactism cannot be the main factor controlling the test building. The main factor is most likely particle shape and mass. In fact the mass is recorded to be roughly proportional to the volume of the particle.
Log-normal grain distributions suggest that the agglutinated testate amoebae produce carefully constructed tests, as also evidenced by grain-size selection and sorting. This log-normal type of distribution implies that larger spaces in the wall between grains should be filled by larger volume of organic cement, rather than an infilling by smaller-sized grains (which would have implied some kind of fractal size distribution, not observed in this study (Tuckwell et al., 1999)). It is more than likely that log-normal grain distributions are the result of a strict exclusion of small grains indicated by the under representation of grains smaller than 0.5–1 μm.
5 Conclusion
Whatever the chemical composition of the sediment, the mean size of the grains agglutinated by testate amoebae is 1.47 ± 0.66 μm. According to this preliminary study, the grain size is a limiting factor for test construction, whereas the mineral composition is not. Hence, when analyzing agglutinated testate amoebae for past reconstructions, it should be taken in account the mean size of the sediment grains that may facilitate or not the construction of the test.
This study calls for further developments, especially whether the sediment grain size may have a significant impact on the species composition of testate amoebae assemblages. The present study was carried out in pristine, oligotrophic environments (mountain lakes with no significant pollutions). Such a study is now to be conducted in environments impacted by ecological stress, for example metal contamination. Do testate amoebae incorporate anthropogenic exogenous particles into their test? If yes, under which conditions?
Acknowledgments
The authors are much indebted to ASTERS society for its help on the project within the Reserve naturelle nationale de Sixt Passy (RNN35) and Fabien Arnaud who helped us with sampling authorizations. The authors are grateful to Emanuela Mattioli and Daniel Gilbert for the thorough suggestions and revisions.