

1 Introduction
Le botulisme est une affection nerveuse commune à l’homme et aux animaux, qui est le plus souvent d’origine alimentaire et qui se traduit par des paralysies flasques. Les intoxications alimentaires sont connues depuis l’antiquité, et le terme botulisme a pour origine le mot latin botulus, désignant une saucisse. Le plus ancien document à ce sujet est un texte indien des XVe–XVIe siècles, qui relate la préparation d’un toxique, probablement de la toxine botulique, à partir de contenu intestinal de mouton. Les premiers cas de botulisme semblent avoir été identifiés par des médecins allemands en 1735. Mais la première description détaillée des symptômes du botulisme a été rapportée par l’Allemand Kerner en 1755 et 1789. Le nombre de cas s’accroît à la fin du XVIIIe–début du XIXe siècle, essentiellement à la suite de la consommation de saucisses. Entre 1820 et 1822, Kerner rapporte 150 cas, dont 80 mortels. Les recherches s’orientent vers la présence de toxiques pouvant se former dans les saucisses ou apportés par les épices et les condiments. Dès 1833, il est noté que l’intoxication par les saucisses en Allemagne est similaire à celle causée par des poissons fumés dans certaines régions de la Russie. Puis, de nombreux cas d’intoxication par des poissons ont été signalés, notamment en Allemagne. La nature bactérienne de cette affection ainsi que la toxine botulique ont été identifiées à la fin du XIXe siècle. Van Ermengem isola pour la première fois en 1897 une bactérie anaérobie toxinogène d’un jambon, ainsi que de l’intestin et de la rate d’une des trois victimes d’un groupe de 30 musiciens qui avait été contaminé 〚3, 38, 59〛. Depuis, de nombreux travaux ont été accomplis et, au cours de ces dix dernières années, des découvertes majeures ont permis de décrypter le support génétique, la structure et le mode d’action au niveau moléculaire des toxines les plus puissantes connues. En effet, la neurotoxine botulique A est la toxine la plus active de toutes les toxines bactériennes, végétales ou animales et de tous les poisons chimiques. La dose létale chez l’homme adulte est estimée entre 100 ng et 1 μg par voie orale 〚46〛. Pour cette raison, la toxine botulique représente une menace d’utilisation comme arme biologique.
2 L’agent du botulisme
Le botulisme résulte de l’action de neurotoxines, dénommées aussi toxines botuliques, qui sont produites par des bactéries anaérobies et sporulées du genre Clostridium.
2.1 Clostridium neurotoxinogènes
Les neurotoxines botuliques, désignées A, B, C1, D, E, F et G, selon leurs propriétés antigéniques, sont produites par diverses espèces de Clostridium. Les bactéries appartenant à ce genre sont des bacilles, anaérobies stricts et sporulés. L’espèce Clostridium botulinum désignait initialement les bactéries productrices d’une toxine induisant une paralysie flasque chez les animaux de laboratoire. Cette espèce est très hétérogène et est divisée en quatre groupes sur la base des propriétés physiologiques, biochimiques et génétiques. En fait, ces quatre groupes correspondent à quatre espèces distinctes sur un plan taxonomique. D’ailleurs, le groupe IV est considéré comme une espèce à part entière, Clostridium argentinense. Il faut noter que chacun des quatre groupes contient des souches non toxinogènes, indifférentiables des souches toxinogènes d’après leurs caractères bactériologiques :
- • groupe I : C. botulinum A et souches protéolytiques de C. botulinum B et F ;
- • groupe II : C. botulinum E et souches glucidolytiques de C. botulinum B et F ;
- • groupe III : C. botulinum C et D ;
- • groupe IV : C. botulinum G ou C. argentinense.
Mais des souches appartenant à d’autres espèces de Clostridium sont également capables de produire une toxine botulique. C’est le cas de certaines souches de Clostridium butyricum, qui synthétisent une neurotoxine botulique E, et de certaines souches de Clostridium baratii, qui produisent une neurotoxine F. Ces souches neurotoxinogènes sont phénotypiquement et génétiquement apparentées aux souches types de C. butyricum et C. baratii, respectivement, et non à celles de C. botulinum.
2.2 Habitat
L’habitat principal des Clostridium neurotoxinogènes, comme des autres espèces de Clostridium, est l’environnement. Grâce à leurs spores, qui sont résistantes aux conditions extrêmes (chaleur, sécheresse, radiation, oxygène, agents chimiques), ces bactéries sont capables de survivre pendant de très longues périodes. Ainsi, les Clostridium sont ubiquistes et largement distribués dans l’environnement. Cependant, la germination des spores et la division bactérienne n’a lieu qu’en conditions anaérobies et en présence de suffisamment de nutriments. Cela restreint l’habitat des Clostridium à des zones anaérobies ou de faible tension en oxygène et contenant des quantités suffisantes de matière organique.
Les différents groupes de C. botulinum n’ont pas une répartition géographique identique. L’habitat principal de C. botulinum A, B, E, F et G est le sol, ainsi que les sédiments marins et d’eau douce. Les toxinotypes A et B sont retrouvés plus volontiers dans les échantillons de sol, mais avec des localisations géographiques différentes. C. botulinum A est prédominant dans la partie ouest des États-Unis, en Amérique du Sud et en Chine, tandis que C. botulinum B est plus fréquent dans la partie est des États-Unis et en Europe. C. botulinum E, qui possède la particularité de se multiplier à basse température (2–3 °C) est préférentiellement retrouvé dans les sédiments marins et d’eau douce, ainsi que dans le contenu intestinal des poissons des régions du Nord de l’hémisphère nord (Alaska, Canada, Scandinavie, Nord de l’Europe, de l’Asie et du Japon). C. botulinum C et D, dont la température optimum de croissance se situe entre 30 et 40 °C et qui sont exigeants en matière organique, sont localisés essentiellement dans des zones riches en matière organique des pays chauds (régions tropicales, régions tempérées en saison chaude). Les cadavres d’animaux morts de botulisme ou porteurs de C. botulinum dans leur tube digestif sont les principaux réservoirs de ces micro-organismes 〚40, 41〛.
C. butyricum et C. baratii sont des bactéries largement répandues dans l’environnement (sol, sédiments, surface de végétaux, végétaux en décomposition, contenu intestinal de l’homme et des animaux sains). La distribution des souches neurotoxinogènes de ces espèces est mal connue.
3 Les toxines botuliques
3.1 Structure des neurotoxines botuliques
Les neurotoxines botuliques sont synthétisées sous la forme d’une seule chaîne protéique de 150 kDa environ, qui est peu ou non toxique. Ces protéines ne possèdent pas de séquence signal, et leur sécrétion intervient par un processus encore mal défini d’exfoliation des parois bactériennes ou par lyse de ces parois. L’activation des neurotoxines fait intervenir une protéolyse, qui clive la protéine précurseur dans son tiers N-terminal en deux fragments dénommés chaîne légère (environ 50 kDa) et chaîne lourde (environ 100 kDa). Les deux chaînes restent réunies par un pont disulfure. Les souches protéolytiques de C. botulinum secrètent une protéase capable d’activer les neurotoxines à l’extérieur de la bactérie. Certaines souches sont non protéolytiques. L’activation peut également survenir sous l’action de protéases d’origine digestives. La trypsine, par exemple, permet d’activer efficacement les neurotoxines botuliques et tétanique. Les neurotoxines botuliques sont associées à d’autres protéines non toxiques pour former des complexes de grande taille (des informations complémentaires sont données en annexe à la fin de ce chapitre).
3.2 Mode d’action
La neurotoxine botulique ingérée ou produite dans la lumière intestinale lors de la toxi-infection traverse la muqueuse intestinale par transcytose, sans causer de lésions. Le rôle dans le franchissement de la barrière digestive des protéines non toxiques formant les complexes botuliques est controversé. Chez la souris, la présence des protéines non toxiques ne serait pas requise pour l’absorption dans la circulation sanguine de la neurotoxine botulique A à partir de l’estomac et de l’intestin 〚32〛. Les composants hemagglutinants de 34 et 50 kDa sont capables de se fixer à la surface des microvilli intestinaux du cobaye et participeraient au transport de la neurotoxine botulique A à travers la muqueuse intestinale 〚21〛. La neurotoxine botulique diffuse dans l’organisme par le biais des circulations sanguine et lymphatique. La mise en évidence de la neurotoxine botulique dans le sérum est la base du diagnostic biologique du botulisme chez l’homme.
Les neurotoxines botuliques ont pour cibles les jonctions neuromusculaires et la neurotoxine tétanique les interneurones inhibiteurs situés dans la moelle épinière. Les deux types de toxines rentrent dans les neurones de façon identique au niveau des extrémités nerveuses démyélinisées. Alors que les toxines botuliques agissent au niveau des jonctions neuromusculaires, la toxine tétanique est véhiculée le long de l’axone jusqu’au corps cellulaire dans la moelle épinière, puis effectue un passage trans-synaptique pour atteindre les interneurones inhibiteurs. Les neurotoxines clostridiennes n’ont pas la possibilité d’atteindre directement les centres nerveux à partir de la circulation sanguine, comme d’autres toxines, en traversant la barrière hématoméningée.
4 Formes de botulisme et modes de contamination
Le botulisme humain est dû aux neurotoxines de type A, B et E produites par C. botulinum et C. butyricum, et exceptionnellement aux types C et F. Le tableau clinique est dominé par une paralysie neuromusculaire, sans anomalie du système nerveux sensitif. Selon le mode d’acquisition, on distingue trois formes de botulisme. En plus des formes naturelles de botulisme, il faut mentionner les risque de botulisme par attentat ou fait de guerre. Étant donné leur activité toxique très élevée, les toxines botuliques représentent des armes biologiques potentielles.
4.1 Intoxination botulique
L’intoxination botulique survient après consommation d’aliments dans lesquels un Clostridium neurotoxinogène s’est développé et a produit des quantités suffisantes de toxine. L’ingestion de toxine préformée dans l’aliment est responsable de l’ensemble des symptômes. C’est la cause la plus commune de botulisme d’origine alimentaire chez l’adulte.
L’incubation est généralement de 12 à 36 h, avec des extrêmes de 5 h à 8 j. Les symptômes débutent le plus souvent par des troubles oculaires, notamment un défaut d’accommodation, mais aussi diplopie, ptôse ou combinaison de ces signes. Puis des dysfonctionnements du système nerveux autonome, tels que sécheresse de la bouche et des yeux, mydriase, difficulté de déglutition, atonie intestinale, constipation, s’installent. Les signes digestifs (nausée, douleurs abdominales, vomissement ou diarrhée) sont présents chez certains patients (environ 50 %) au tout début de l’affection (24 premières heures), puis disparaissent. Ils pourraient être dus à d’autres agents et toxines, responsables d’intoxications alimentaires. Les formes plus graves se traduisent par une faiblesse des membres et des difficultés respiratoires. La mort peut survenir par insuffisance respiratoire 〚45, 53〛.
4.2 Toxi-infection botulique
L’ingestion de spores ou de formes végétatives de Clostridium neurotoxinogènes peut, dans certaines circonstances, s’accompagner d’une multiplication bactérienne et de production de toxine dans le contenu intestinal. Habituellement, la flore digestive résidente prévient la colonisation de l’intestin par une bactérie étrangère apportée par l’alimentation. Chez les jeunes enfants, une flore digestive incomplètement constituée ou incomplètement fonctionnelle peut permettre l’implantation de Clostridium neurotoxinogènes dans l’intestin : c’est le botulisme infantile. Selon diverses enquêtes épidémiologiques, une dizaine de spores de C. botulinum est suffisante pour déclencher la maladie chez un jeune enfant. Les Clostridium neurotoxinogènes et la toxine sont retrouvés dans le contenu intestinal ; la toxine est rarement identifiée dans le sérum, contrairement à l’intoxination botulique, dans laquelle la toxémie est fréquente.
La plupart des souches impliquées dans le botulisme infantile appartiennent au groupe I (C. botulinum A et souches protéolytiques de C. botulinum B). Les souches de C. botulinum A sont divisées en deux sous-groupes, dénommés A1 et A2, selon le polymorphisme des profils de restriction d’ADN. Les souches de botulisme infantile isolées au Japon appartiennent au sous-groupe A2, les souches isolées de botulisme chez l’adulte et les nouveau-nés aux États-Unis se répartissent dans les sous-groupes A1 et A2 〚7〛.
La toxi-infection botulique survient aussi chez les adultes. Les facteurs prédisposants, tels que chirurgie intestinale, antibiothérapie, inflammation chronique ou lésions chroniques de la muqueuse intestinale, permettraient la croissance de Clostridium neurotoxinogènes dans le tube digestif.
4.3 Botulisme par blessure
Les plaies peuvent être colonisées par C. botulinum et provoquer l’apparition d’un botulisme. Ce mode de contamination est analogue à celui du tétanos ou d’une gangrène à Clostridium. Les plaies anfractueuses, profondes et avec une faible ouverture sur l’extérieur, sont favorables au développement de bactéries anaérobies. La présence de tissu nécrosé, suite à une contusion par exemple, stimule la croissance des Clostridium, en apportant les nutriments nécessaires à ces bactéries, qui n’envahissent pas les cellules saines. Les C. botulinum responsables du botulisme par blessure appartiennent aux toxinotypes A et B du groupe I. Cependant, l’incidence du botulisme par blessure est beaucoup moins fréquente que celle du tétanos.
Le botulisme par blessure est en recrudescence aux États-Unis et en Europe chez les toxicomanes, soit par l’utilisation de matériel contaminé lors des injections, soit du fait de la présence de spores de C. botulinum dans les drogues 〚48, 55〛.
4.4 Situation du botulisme humain en France
En France, le botulisme est une maladie à déclaration obligatoire de façon séparée des autres toxi-infections alimentaires depuis 1986. Le nombre de cas annuels varie de 10 à 27. Le type le plus représenté est le type B, traditionnellement à la suite d’une consommation de jambon salé et non cuit ou de charcuteries familiales. Il est à remarquer que le nombre de cas de botulisme est resté pratiquement constant au cours de ces 30 dernières années, alors que les habitudes alimentaires ont sensiblement changé. La consommation de conserves familiales a régressé au profit de celle d’aliments ou de préparations culinaires d’origine commerciale et de la prise plus fréquente de repas dans des restaurants (restaurants d’entreprise ou autres). L’investigation de l’origine de ces nouvelles formes de botulisme est plus difficile à mettre en œuvre. Un cas récent de botulisme de type A avait pour origine une soupe industrielle de poisson. L’interrogatoire des patients révèle de plus en plus souvent qu’ils n’avaient pas consommé de conserves familiales et qu’ils avaient, soit consommé des aliments du commerce sans les avoir stockés sur une longue période, soit pris leurs repas dans des restaurants. Les nouvelles formes de conserves d’aliments (emballage sous vide, aliments frais ou pasteurisés et conservés réfrigérés) sont propices au développement de Clostridum neurotoxinogènes. Les récents cas de botulisme E n’ont pas tous été élucidés. Dans un cas, une marinade de poisson a été suspectée d’en être à l’origine ; dans les autres cas, les personnes concernées ont affirmé avoir fréquenté des restaurants au cours des jours qui ont précédé l’apparition des symptômes.
5 Neurotoxines botuliques en tant qu’armes biologiques
Étant donné leur activité biologique élevée, les neurotoxines botuliques représentent des armes potentiellement très dangereuses. Elles pourraient être utilisées pour contaminer une alimentation en eau potable, une usine de mise en bouteille d’eau ou d’une autre boisson, une chaîne de préparation d’aliments ou de repas cuisinés à l’avance.
Un exemple de bioterrorisme classiquement évoqué est celui de la contamination d’un réseau de distribution d’eau avec de la toxine botulique. Les effets escomptés d’un tel acte en terme de personnes atteintes et de mortalité sont fonction de nombreux paramètres : quantité de toxine inoculée, site d’inoculation dans le réseau, complexité ou maillage du réseau... De plus, la diffusion généralisée de la toxine dans le réseau est difficilement prévisible. Étant donné qu’une faible proportion seulement de l’eau utilisée est ingérée, il n’est pas certain qu’une telle opération atteigne un grand nombre de personnes.
Cependant, la répercussion médiatique, avec affolement de la population, mise en place d’une infrastructure de substitution d’approvisionnement en eau (interdiction d’utilisation de l’eau du robinet, distribution d’eau en bouteille ou par citerne...) et nécessité de décontamination du réseau, serait incalculable.
Les principales mesures préventives comprennent : la surveillance des accès aux sites sensibles du réseau de distribution d’eau (réservoirs, principales canalisations, usines de traitement...), la détection d’introduction de substances étrangères dans le réseau (en utilisant par exemple les capteurs de chlore), l’identification spécifique de la toxine botulique et la chloration de l’eau (la toxine botulique est très sensible aux agents oxydants).
6 Identification
L’identification du botulisme repose, d’une part, sur le diagnostic de cette affection chez le patient et, d’autre part, sur l’investigation de l’aliment à l’origine de la contamination.
Les signes cliniques de paralysie flasque avec atteinte oculaire précoce sont évocateurs de botulisme, mais le diagnostic différentiel doit être fait avec d’autres affections nerveuses.
L’intoxination botulique s’accompagne fréquemment chez l’homme d’une toxémie, c’est-à-dire de la présence de toxine botulique dans la circulation sanguine. La mise en évidence et le typage de la toxine botulique dans le sérum des patients est une méthode fiable de diagnostic du botulisme. Le test le plus utilisé est le test de létalité sur souris, car il est plus sensible que les méthodes immunologiques de type Elisa 〚12, 13, 35, 49〛. Les titres en toxine botulique dans le sérum des malades sont de l’ordre de 0,5 à 4 doses létales souris par millilitre. Des tests basés sur l’activité biochimique, c’est-à-dire sur l’activité protéasique des neurotoxines botuliques vis-à-vis de peptides spécifiques de synaptobrévine, SNAP25 ou syntaxine, sont en cours de développement, notamment pour la détection de la neurotoxine botulique B 〚1, 60〛.
Dans les formes de toxi-infections de botulisme, le Clostridium neurotoxinogène et la toxine botulique sont présents dans le contenu intestinal et les selles du patient. Il faut noter que, dans ces formes, notamment dans le botulisme infantile, la toxine botulique est rarement présente dans le sérum. La recherche de Clostridium neurotoxinogène et de toxine botulique dans les selles sont les éléments du diagnostic biologique de ces formes de botulisme. L’isolement et l’identification des Clostridium neurotoxinogènes par les méthodes standard de bactériologie sont complexes et peu fiables, du fait que ces bactéries sont des anaérobies stricts, avec des caractères phénotypiques variés, et qu’il n’existe pas de milieu sélectif efficace. Actuellement, la recherche de Clostridium neurotoxinogène est réalisée par amplification génique (PCR) d’un fragment d’ADN de gène de neurotoxine à partir d’une culture d’enrichissement de l’échantillon. Plusieurs méthodes de biologie moléculaire ont été proposées pour la détection des Clostridium botulinum et Clostridium neurotoxinogènes apparentés 〚2, 4, 6, 7, 16–19, 27, 29, 50–52, 54〛.
La recherche de toxine botulique et/ou de Clostridium neurotoxinogène dans les aliments est réalisée avec les mêmes méthodes que celles utilisées pour les échantillons biologiques. Il faut noter la nécessité de disposer d’un test rapide de détection de la neurotoxine botulique dans des échantillons alimentaires ou d’eau. Un test immunologique sur bandelette est proposé par l’armée américaine pour le dépistage de la toxine botulique A, mais sa sensibilité est très inférieure à celle du test sur souris.
7 Traitement
Le traitement du botulisme est le plus souvent symptomatique. Il consiste essentiellement en une assistance respiratoire, qui, dans les cas graves, comprend trachéotomie et réanimation, d’une durée allant de plusieurs jours à plusieurs semaines.
Le traitement spécifique repose sur la sérothérapie, qui consiste en l’administration d’anticorps préparés chez le cheval contre les toxines de type A, B et E. Ces sérums trivalents ne sont plus actuellement préparés en Europe, et leur obtention auprès du Center of Disease Control d’Atlanta reste limitée. L’efficacité de la sérothérapie est discutée et aucune étude prospective n’a été réalisée. L’injection d’anticorps anti-toxine botulique a pour effet de neutraliser la toxine botulique circulante, mais elle est inopérante sur celle liée de façon irréversible au tissu nerveux ou celle internalisée dans les neurones. La demi-vie des anticorps injectés serait de 5 à 8 j. De ce fait, la sérothérapie ne peut être efficace que si elle est instaurée précocement après l’apparition des symptômes. Ainsi, dans une analyse de 134 cas de botulisme de type A, la mortalité était de 10 % chez les patients ayant reçu une sérothérapie dans les 24 premières heures, de 15 % chez ceux traités tardivement et de 46 % chez ceux qui n’avaient pas bénéficié de ce traitement. La sérothérapie a pour principal inconvénient d’induire chez certaines personnes (jusqu’à 9 %) une réponse allergique 〚48, 53〛. Actuellement, les sérums anti-botuliques ne sont plus, ni produits, ni commercialisés par les firmes pharmaceutiques. Des réserves d’anticorps anti-botuliques sont à disposition sur le continent Nord-Américain (Center of Disease Control) et en quantité très limitée en Europe. Les autorités de défense se préoccupent actuellement de reconstituer des stocks de sérums ou d’anticorps purifiés anti-neurotoxines botuliques, pour parer à d’éventuels attentats biologiques. Dans le cadre du traitement du botulisme infantile, qui est particulièrement fréquent aux États-Unis, des anticorps humains anti-neurotoxine botulique A, B et E ont été proposés 〚20〛.
L’antibiothérapie (notamment pénicillines) est recommandée dans les toxi-infections botuliques comme le botulisme infantile et le botulisme par blessure. Son efficacité est incertaine dans le botulisme d’origine alimentaire dû à une ingestion de toxine préformée.
Les composés capables de stimuler la libération d’acétylcholine aux jonctions neuromusculaires ont été largement évalués dans le traitement du botulisme. La guanidine, comme les aminopyridines, prolongerait le potentiel d’action au niveau des terminaisons nerveuses en bloquant les canaux potassiques et, par voie de conséquence, en stimulant les canaux calciques et en facilitant l’entrée du calcium. Ces drogues pourraient aussi avoir un effet sur la mobilité du calcium intracellulaire. La guanidine s’est montrée efficace pour restaurer la transmission nerveuse des préparations in vitro nerf–muscles d’animaux expérimentalement intoxiqués par la toxine botulique A. Les effets bénéfiques de la guanidine chez les sujets atteints de botulisme A ou B étaient variables. Certains patients avaient montré une légère amélioration transitoire 〚53〛.
Les aminopyridines, notamment la 4-aminopyridine et la 3,4-diaminopyridine, sont 20 à 30 fois plus actives que la guanidine. Ces composés restaurent l’activité motrice d’animaux intoxiqués par la toxine botulique A, mais ils n’ont qu’un faible effet sur les autres types de toxines botuliques. La spécificité des aminopyridines pour le type A n’est pas connue. Il faut noter que ces drogues ont des effets indésirables, comme aplasie médullaire, néphrite, arythmie, dysfonctionnement hépatique, etc., pour la guanidine, et convulsions, pour les aminopyridines 〚53〛.
8 Immuno-prévention
La prévention médicale du botulisme est basée essentiellement sur la vaccination. La vaccination anti-botulique est efficace pour prévenir le botulisme de façon comparable à celle du tétanos. Cependant, les vaccins actuels, préparés à partir de préparations brutes de neurotoxines botuliques, se révèlent parfois allergisants. La vaccination anti-botulique de masse ne se justifie pas pour prévenir le botulisme. Elle est réservée aux personnes à risque, personnel de laboratoire ou militaires devant intervenir dans des conflits où il existe une possibilité d’utilisation d’armes biologiques. Par ailleurs, les neurotoxines botuliques sont utilisées en thérapeutique pour traiter des paralysies spastiques permanentes (blépharospasme, torticolis rebelle, paralysie hémifaciale, crampe de l’écrivain...). Ainsi, une vaccination anti-botulique généralisée ne permettrait pas aux patients de bénéficier d’un traitement par la neurotoxine botulique.
Une nouvelle forme de vaccins est en cours de développement. Il s’agit de vaccins génétiquement modifiés, qui sont préparés à partir de neurotoxine recombinante mutée. La protéine recombinante correspond, soit à la neurotoxine entière, avec des mutations ponctuelles sur le site actif, soit à un domaine de la neurotoxine qui est non toxique et qui stimule la production d’anticorps neutralisants. Il a été montré que le domaine Fc (moitié C-terminale de la chaîne lourde qui constitue le domaine de reconnaissance du récepteur à la surface des neurones) est aussi efficace que l’anatoxine pour induire une réponse immunologique protectrice. D’importants travaux ont été accomplis, notamment par l’armée américaine, pour produire des vaccins recombinants à partir des fragments Fc des différents types de neurotoxines botuliques 〚5〛. Les fragments Fc étant plus courts que la neurotoxine entière, ils sont plus facilement produits comme protéine recombinante.
9 Nouvelles approches thérapeutiques
Les neurotoxines botuliques et tétaniques sont des métallo-protéases à zinc. Une possibilité consiste à utiliser des inhibiteurs de protéases, à condition que ces agents puissent être transportés dans les cellules nerveuses et n’interfèrent pas avec des protéases endogènes ou d’autres composés de la cellule. Les inhibiteurs classiques (phosphoramidon, captopril) des métalloprotéases à zinc et les autres inhibiteurs de protéases se sont révélés inopérants. Des inhibiteurs tripeptidiques de la toxine tétanique et de la neurotoxine botulique B ont été développés. Ces inhibiteurs sont spécifiques de chaque toxine. Ils agissent en bloquant le site enzymatique et ont une activité de l’ordre du micromolaire et nanomolaire respectivement 〚33〛 (C. Anne et B. Roques, non publié).
Une autre approche consiste à développer une stratégie d’internalisation d’anticorps neutralisants des neurotoxines botuliques dans les cellules cibles. Ainsi, ces anticorps pourraient neutraliser les neurotoxines à l’intérieur des neurones. Les toxines ont la propriété de pénétrer efficacement dans les cellules et peuvent être utilisées pour véhiculer des protéines hétérologues dans les cellules. Les méthodes de la biologie moléculaire permettent de modifier les toxines au niveau leur activité enzymatique ou de leur site de liaison à des récepteurs cellulaires. Ainsi, le domaine de reconnaissance de récepteur à la surface des cellules peut être interchangé avec un domaine interagissant spécifiquement avec un récepteur de neurone. Par exemple, il est envisageable de coupler le domaine fonctionnel d’un anticorps anti-neurotoxine (fragment variable simple chaîne) avec la chaîne lourde d’une neurotoxine botulique, qui assure l’internalisation spécifique dans les neurones et qui est dépourvue d’activité toxique. Un autre modèle toxinique peut être également sélectionné, dans lequel le domaine enzymatique est remplacé par le domaine fonctionnel d’un anticorps, et celui de reconnaissance de récepteur cellulaire interchangé avec un autre, spécifique des neurones. Des travaux sont en cours, pour valider un tel modèle, avec une toxine binaire de Clostridium perfringens.
Annexe
A1 Structure
Les neurotoxines botuliques sont associées à d’autres protéines non toxiques pour former des complexes de grande taille. Ces protéines incluent une protéine, appelée non toxique non hémaglutinine (NTNH), qui a une taille proche de celle des neurotoxines (139 kDa), et des protéines qui ont une activité hémaglutinante (HA), dont les tailles sont 34, 17 et 70 kDa chez C. botulinum A. La neurotoxine s’associe à NTNH pour former des complexes de taille moyenne (300 kDa) (M) et leur association aux HA est à l’origine des complexes de grande taille (500 kDa) (L). Des complexes de très grande taille (900 kDa) (LL), observés notamment chez C. botulinum A, résultent d’une dimérisation des complexes L. Les protéines NTNH sont très conservées au niveau de leur séquence en acides aminés (70–80 % d’identité) et elles présentent peu d’homologie avec les neurotoxines botuliques (31–39 % d’identité). Toutefois, les 100 premiers acides aminés des protéines NTNH sont les plus apparentés aux acides aminés correspondants des neurotoxines botuliques, ce qui pourrait rendre compte de la liaison NTNH–neurotoxine. Le rôle des complexes est encore mal compris. Ils pourraient protéger les neurotoxines botuliques vis-à-vis des conditions dénaturantes, telles que l’acidité de l’estomac et les protéases digestives. Il est à noter que la toxine tétanique ne s’associe pas à d’autres protéines pour former des complexes et qu’elle n’est pas stable dans le tractus digestif. Les protéines non toxiques pourraient intervenir dans le passage de la barrière intestinale.
Les gènes des neurotoxines clostridiennes et des protéines des complexes botuliques ont été caractérisés (pour une revue, voir 〚41〛). Ils sont à proximité l’un de l’autre dans une même région d’ADN, dénommée locus botulique. Ils sont organisés en deux opérons, l’un comprenant les gènes codant pour NTNH et la neurotoxine, l’autre les gènes des composants hémaglutinines. Un gène régulateur positif (botR) de l’expression de ces gènes a une position variable chez C. botulinum. Il est situé, soit en partie 5’ du locus botulique chez C. botulinum C et D, soit entre les deux opérons chez C. botulinum A, B, E et F. C’est le seul gène du complexe botulique qui est conservé chez C. tetani. Le gène tetR, homologue de botR, est localisé en amont du gène de la neurotoxine tétanique. Les gènes des protéines non toxiques des complexes botuliques sont absents chez C. tetani. Les gènes des neurotoxines et des protéines non toxiques associées aux neurotoxines botuliques ont des localisations diverses : chromosomique chez C. botulinum A, B, E et F, plasmidique chez C. botulinum G et C. tetani, et phagique chez C. botulinum C et D. De plus, il semble que les locus botuliques soient situés sur des éléments génétiques mobiles, tels que des transposons. Cela rendrait compte de la mobilité des gènes des neurotoxines entre les diverses structures d’ADN et les échanges entre souches. Ainsi, dans chaque type de C. botulinum et chez C. tetani, il existe des souches variantes qui ont perdu leur toxinogenèse et qui peuvent redevenir toxiques par acquisition des gènes des neurotoxines au moyen de transfert de phage, de plasmide ou de transposon par conjugaison. Certaines souches possèdent plusieurs gènes de neurotoxines ou produisent des toxines hybrides. Des toxines mosaïques entre les types C et D ont été décrites. Par ailleurs, certaines souches d’espèces de Clostridium, habituellement non toxinogènes, sont productrices de neurotoxines botuliques et sont responsables du botulisme chez l’homme. C’est le cas des souches de Clostridium butyricum, qui produisent une neurotoxine botulique de type E, et des souches de Clostridium baratii, qui synthétisent une neurotoxine de type F.
Les longueurs des séquences en acides aminés des neurotoxines botuliques et tétanique varient de 1251 à 1315. La plus longue séquence est celle de la neurotoxine tétanique. Les homologies sont comprises entre 34 à 97 % d’identité. La neurotoxine botulique B est celle qui est le plus apparentée à la neurotoxine tétanique (42 % d’identité). L’analyse génétique a révélé une grande diversité parmi les neurotoxines botuliques (au moins 11 variants), plus importante que les sept toxinotypes classiques, alors qu’un seul type de neurotoxine tétanique est connu. Les régions les plus conservées concernent les 100 premiers acides aminés et la région centrale de la chaîne légère, ainsi que la moitié N-terminale de la chaîne lourde. La région centrale des chaînes légères contient un motif (HExxH) caractéristique des métalloprotéases dépendantes du zinc. L’identification de ce motif chez la neurotoxine tétanique, qui fut la première à être séquencée, fournit la première indication que les neurotoxines clostridiennes sont des métalloprotéases. Il fut ensuite démontré que chaque chaîne légère lie un atome de zinc et que les agents chélateurs du zinc inhibent l’activité des neurotoxines. La moitié N-terminale de la chaîne lourde est impliquée dans la translocation de la chaîne légère dans le cytosol. Le degré élève de conservation de ce domaine suppose que toutes les neurotoxines suivent un même processus d’entrée dans les neurones. En revanche, chaque neurotoxine reconnaît un récepteur distinct en surface des neurones. Le faible niveau d’identité de la moitié C-terminale des chaînes lourdes, qui est impliquée dans la liaison au récepteur, reflètent cette divergence 〚47〛.
Les structures tridimensionnelles des neurotoxines botuliques A et B ainsi que de la partie C-terminale de la toxine tétanique ont été déterminées. Cette structure révèle trois domaines distincts de 50 kDa environ, qui sont en accord avec les trois domaines fonctionnels définis auparavant par des méthodes biochimiques et physiologiques : la chaîne légère, qui comporte le site enzymatique, la moitié N-terminale de la chaîne lourde, qui correspond au domaine de translocation, et la moitié C-terminale, qui est le domaine interagissant avec le récepteur à la surface des neurones.
Les structures du domaine C-terminal des chaînes lourdes des neurotoxines tétanique et botulique A sont très similaires. Il est divisé en deux sous-domaines, formés de feuillets bêta. L’un en C-terminal est homologue à d’autres protéines ou domaines ayant des fonctions de liaison à un récepteur. Il serait impliqué dans la reconnaissance d’une glycoprotéine, alors que celui en amont, qui est relativement bien conservé parmi toutes les neurotoxines, reconnaîtrait un ganglioside. Le domaine N-terminal de la chaîne lourde de la neurotoxine botulique A a une forme cylindrique et contient deux hélices alpha inhabituellement longues, qui entourent la chaîne légère et plusieurs courtes hélices alpha. Cette structure ressemble à celle de protéines virales qui changent de conformation à pH acide. La chaîne légère possède des hélices alpha et des feuillets bêta. Elle est peu apparentée à d’autres enzymes de structure connue, à l’exception du motif liant le zinc. La coordination du zinc par les deux His du motif HExxH par les résidus Glu-261, Tyr-365 et une molécule d’eau liée à la Glu du motif présente certaines homologies avec celle de la thermolysine, mais possède une architecture distincte. Le site actif est masqué par les hélices du domaine N-terminal de la chaîne lourde. De ce fait, la neurotoxine entière est dépourvue d’activité enzymatique, celle-ci est exercée par la chaîne légère dissociée de la chaîne lourde.
A2 Mode d’action
Le mode d’action moléculaire des neurotoxines clostridiennes, qui a pour effet le blocage de la libération des neuromédiateurs par les neurones intoxiqués, fait intervenir quatre étapes : liaison à un récepteur à l’extrémité démyélinisée de neurones, internalisation, translocation à travers la membrane des vésicules d’endocytose et modification enzymatique d’une cible intracellulaire 〚10, 11, 24, 42, 47〛.
A3 Liaison au récepteur
Après diffusion par le sang et/ou la lymphe, les neurotoxines botuliques se fixent sur les extrémités démyélinisées des motoneurones (Fig. 1). La neurotoxine tétanique se lie également aux motoneurones, mais aussi aux fibres sensitives et adrénergiques. La nature du récepteur des neurotoxines est encore controversée. De nombreux travaux ont montré que les neurotoxines se lient à des gangliosides, tels que GD1B, GT1B et GQ1B. Cependant, cette interaction est de faible affinité et ne rend pas compte de la spécificité de reconnaissance de chaque neurotoxine. Un modèle de double récepteur, comprenant une glycoprotéine et un ganglioside, a été proposé à partir d’études de liaison des neurotoxines sur cultures cellulaires et d’après la structure du domaine de liaison au récepteur en deux sous-domaines. Cela est conforté par le fait que la neurotoxine botulique B se fixe avec une forte affinité à la synaptotagmine II des membranes des vésicules synaptiques en présence de ganglioside. Des isoformes de synaptotagmines seraient également les récepteurs des neurotoxines botuliques A et E. Celui de la neurotoxine tétanique n’a pas encore été identifié. Sa partie protéique correspondrait à une protéine membranaire de 15 kDa, qui aurait un rôle déterminant dans le routage de la toxine 〚25〛. Cette protéine, qui serait apparentée à la protéine Thy1, est liée à un glycosylphosphatidylinositol et est localisée sur un microdomaine lipidique à la surface des cellules neuronales. La toxine tétanique se lie à une récepteur situé sur des radeaux lipidiques, et le cholestérol est important pour l’internalisation et/ou le trafic intracellulaire de la toxine dans les neurones 〚26〛.

Trajet des neurotoxines botuliques et de la toxine tétanique jusqu’aux terminaisons nerveuses.
A4 Internalisation
Les neurotoxines clostridiales ne traversent pas directement la membrane cytoplasmique. Elles sont internalisées par endocytose dans des compartiments intracellulaires acides. Il a été supposé que la neurotoxine tétanique serait internalisée dans des vésicules synaptiques, au cours du recyclage exocytose/endocytose. La majorité des vésicules synaptiques, après leur phase d’exocytose libérant le neuromédiateur dans la fente synaptique, est recyclée par endocytose et rechargée en neuromédiateur. Cela est en accord avec l’observation du fait que la stimulation nerveuse facilite l’intoxication. Mais ces résultats sont actuellement controversés ; en effet, la toxine tétanique est capable d’entrer dans des neurones intoxiqués par la toxine botulique A, c’est-à-dire quand l’exocytose est bloquée 〚24〛. L’identité des vésicules permettant l’entrée des neurotoxines botuliques n’est pas encore connue. Il est probable que les neurotoxines botuliques et tétanique entrent dans des vésicules différentes. Une grande différence entre les deux sortes de neurotoxines est que les neurotoxines botuliques restent à l’extrémité présynaptique des motoneurones, alors que les vésicules contenant la neurotoxine tétanique suivent un transport rétrograde le long de l’axone jusqu’aux corps cellulaires dans la mœlle épinière.
A5 Translocation dans le cytosol
Quelles que soient les vésicules d’endocytose contenant les neurotoxines, la chaîne légère doit traverser la membrane vésiculaire pour se localiser dans le cytosol, où elle exerce son activité. L’acidification des vésicules au moyen de l’ATP-ase vésiculaire est une étape indispensable pour la translocation. La baisse de pH induit un changement de conformation d’une forme hydrophile à pH neutre à une forme dont les segments hydrophobes sont exposés en surface à pH acide, permettant l’insertion de l’ensemble de la neurotoxine dans la bicouche lipidique. L’acidification induit l’oligomerisation des chaînes lourdes de neurotoxine en tetramères, qui s’insèrent dans la bicouche lipidique et forment des canaux ioniques. Ces tetramères résultent de l’association d’hélices amphiphiles localisées dans le domaine N-terminal des chaînes lourdes.
Le mécanisme de translocation est encore hypothétique. Selon un premier modèle, la chaîne légère, dépliée à pH acide, passerait à travers le canal formé par les chaînes lourdes oligomérisées. L’exposition au pH neutre dans le cytosol permettrait le repliement de la chaîne légère et sa dissociation de la vésicule par réduction du pont disulfure entre les deux chaînes de neurotoxine. Cependant, les valeurs expérimentales de la taille des pores sont incompatibles avec le passage d’une molécule de la taille de la chaîne légère. Les canaux formés par les chaînes lourdes pourraient altérer le gradient électrochimique et induire une lyse osmotique permettant la traversée de la neurotoxine. Mais ce phénomène de lyse cellulaire n’a pas été observé expérimentalement. Un troisième modèle est celui du sillon. Les chaînes lourdes insérées dans la membrane formeraient un sillon hydrophile. La chaîne légère, partiellement dépliée, s’engagerait dans ce sillon, sa partie hydrophile du côté des chaînes lourdes et sa partie hydrophobe faisant face aux lipides membranaires.
A6 Activité enzymatique
Les neurotoxines clostridiennes sont des protéases d’une remarquable spécificité. Cette activité est médiée par la chaîne légère, qui contient dans sa région centrale un motif conservé (HExxH), qui correspond au site enzymatique. Les substrats des neurotoxines clostridiennes sont trois protéines, dénommées protéines SNAREs, qui comprennent la VAMP (Vesicle Associated Membrane Protein) ou synaptobrevine, la SNAP25 et la syntaxine. La neurotoxine tétanique et les neurotoxines botuliques B, D, F et G clivent la protéine VAMP, les neurotoxines botuliques A et E clivent SNAP25 et la neurotoxine botulique C1 clive à la fois la syntaxine et SNAP25. Les sites de clivage sont différents pour chaque neurotoxine, sauf pour la neurotoxine tétanique et la neurotoxine botulique B. Ces deux neurotoxines ont le même substrat, la protéine VAMP, qu’elles coupent au même site ; pourtant, elles causent des tableaux cliniques opposés, à savoir le tétanos et le botulisme, respectivement. Cela souligne le fait que les symptômes différents observés résultent du site de l’intoxication plutôt que du mécanisme moléculaire d’action des deux toxines.
VAMP est une protéine de 13 kDa, qui est ancrée dans la membrane des vésicules synaptiques. Elle possède quatre domaines : un segment variable riche en proline, correspondant aux 33 acides N-terminaux, suivi d’un segment conservé, formant une structure enroulée, d’un domaine transmembranaire et d’une extrémité conservée intravésiculaire. Dix isoformes ont été identifiées à ce jour et sont distribuées dans divers tissus, certaines étant plus spécifiques du tissu nerveux (VAMP1, VAMP2) et d’autres plus ubiquitaires (VAMP3 ou cellubrevine). Certaines sont clivables par les neurotoxines clostridiennes, d’autres sont résistantes (TI-VAMP), du fait de mutations dans le site de clivage et les sites de liaison aux neurotoxines 〚22〛. Cela rend compte, du moins en partie, de la résistance ou de la sensibilité de certaines espèces animales au tétanos et au botulisme. Ainsi, l’isoforme 1 de VAMP du rat et du poulet n’est pas clivable par la neurotoxine tétanique et la neurotoxine botulique B du fait d’une mutation (Gln-Val) au niveau du site de clivage. Or, le rat et le poulet présentent une résistance élevée à la neurotoxine tétanique par rapport à l’homme et à la souris chez lesquels VAMP1 est clivable.
Dans les vésicules synaptiques, VAMP est associée à d’autres protéines, telles que synaptophysine, l’ATPase vésiculaire, un accepteur pour la protéine Rab. VAMP2 interagit avec la protéine adaptatrice AP3 et aurait un rôle dans la formation du manteau des vésicules synaptiques.
SNAP25 est palmytoilée au niveau d’une cystéine localisée au milieu de la molécule, permettant ainsi son insertion dans la membrane présynaptique. SNAP25 forme un complexe avec la synaptotagmine, qui est une protéine Ca++-senseur et jouerait un rôle important dans la neurotransmission dépendante du calcium. Les deux isoformes A et B de SNAP25 sont localisées dans les neurones. L’isoforme SNAP23 est exprimée dans les cellules non neuronales et est impliquée dans l’exocytose. SNAP23 de souris est clivable par la neurotoxine botulique A et E, tandis que SNAP23 d’origine humaine n’est pas clivable 〚22〛.
La syntaxine (35 kDa) est localisée principalement dans la membrane présynaptique, en association avec différents types de canaux calciques. Le domaine N-terminal est constitué de trois longues hélices alpha, qui sont impliquées dans des interactions protéine–protéine. Un domaine transmembranaire est situé en C-terminal. La famille des syntaxines regroupe au moins 20 isoformes, localisées dans le tissu nerveux et aussi au niveau des granules chromaffines. Certaines isoformes telle syntaxine 3 sont non clivables par la neurotoxine botulique C1 〚22〛.
La syntaxine, SNAP25 et VAMP s’associent pour former un complexe (complexe SNARE), qui joue un rôle essentiel dans la neuroexocytose. La structure de ce complexe a été récemment élucidée. Il est formé de quatre hélices alpha, enroulées parallèlement les unes sur les autres et provenant de l’association de la majeure partie de domaines cytoplasmiques de VAMP, syntaxine et des segments N- et C- terminaux de SNAP25. Cette dernière protéine contribue à la formation de deux hélices alpha parallèles. Les protéines restent associées aux membranes vésiculaire et présynaptique. Ces enroulements ont pour effet de rapprocher fortement les deux types de membrane, les complexes SNAREs étant dirigés parallèlement à la membrane. Le modèle SNARE propose que des protéines de la membrane vésiculaire (vSNAREs) s’associent à des protéines d’une membrane cible (tSNAREs), de façon à former un complexe permettant l’accrochage des deux types de membrane et ensuite leur fusion.
Les complexes SNAREs (coefficient de sédimentation 7 s) ainsi formés sont stables et ont la possibilité de recruter d’autres protéines cytosoliques, pour former un complexe de plus grande taille (complexe 20 s). Les facteurs cytosoliques sont le N-ethylmaleimide-sensitive factor (NSF) et les NSF attachment proteins (αSNAPs). NSF est une ATPase qui a été initialement identifiée comme une protéine essentielle dans le transport vésiculaire entre les différents compartiments de l’appareil de Golgi. En présence d’ATP, NSF catalyse la dissociation des complexes SNAREs en leurs monomères.
A7 Reconnaissance spécifique des cibles par les neurotoxines clostridiennes
Les cibles (VAMP, SNAP25 et syntaxine) ne présentent pas de séquence conservée en acides aminés au niveau et dans la proche région des sites de clivage par les neurotoxines. De plus, des peptides synthétiques courts comprenant les sites de clivage ne sont pas protéolysés par les neurotoxines clostridiennes. Seuls des peptides longs (40 acides aminés et plus) sont clivés. Cela suggère que les neurotoxines reconnaissent principalement la structure tertiaire de leurs cibles. Un motif de neuf acides aminés, adoptant une structure en hélice alpha avec trois résidus chargés négativement sur une face et trois résidus hydrophobes sur l’autre face, est conservé parmi les trois cibles des neurotoxines. Ce motif, appelé motif SNARE, existe en deux copies chez VAMP (V1 et V2) et syntaxine (X1 et X2) et en quatre copies chez SNAP25 (S1, S2, S3 et S4). Il a été montré expérimentalement que ce motif est impliqué dans la liaison avec les neurotoxines. Ainsi, les peptides synthétiques incluant le site de clivage et ces motifs sont efficacement protéolysés par les neurotoxines. De plus, la neurotoxine tétanique dont la cible est VAMP reconnaît le motif V1 et un motif chargé positivement 〚47〛. Ces deux motifs sont situés de part et d’autre du site de clivage. La neurotoxine tétanique se comporterait comme une enzyme allostérique et serait activée par changement de conformation après liaison à ces deux motifs de VAMP 〚8〛.
Les régions des neurotoxines tétanique et botuliques impliquées dans la liaison avec le substrat ne sont pas connues.
A8 Inhibition de la neuroexocytose
Les neurotoxines clostridiennes agissent en inhibant la sécrétion des neuromédiateurs. Puisque les neurotoxines clostridiennes sont des métalloprotéases dont les substrats sont VAMP, SNAP25 et syntaxine, ces trois protéines jouent un rôle essentiel dans la neuroexocytose. Il faut noter que les neurotoxines clivent leurs substrats quand ils sont sous formes monomériques et non quand ils sont associés dans le complexe SNARE. Le modèle proposé du mécanisme de neurosécrétion est le suivant.
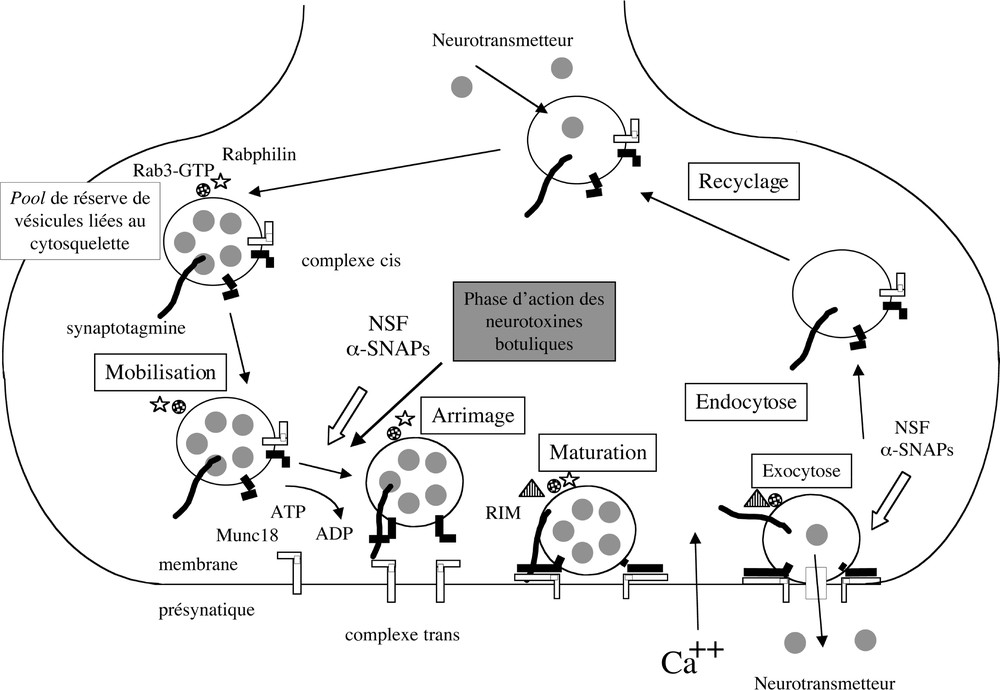
Dans les terminaisons nerveuses, le neuromédiateur (acétylcholine pour les motoneurones) s’accumule dans les vésicules synaptiques par l’intermédiaire de transporteurs spécifiques et par un pH de gradient généré par la pompe à proton, l’ATPase vésiculaire. Le cycle des vésicules synaptiques comprend la mobilisation et l’amarrage à la membrane présynaptique (docking), puis une phase d’apprêtage (priming), permettant aux vésicules d’effectuer l’étape de fusion membranaire et d’exocytose du neuromédiateur, en réponse à une entrée cellulaire d’ions calcium. Les vésicules synaptiques sont ensuite recyclées par endocytose.
Les vésicules synaptiques sont liées aux filaments d’actine par l’intermédiaire de la synapsine et sont mobilisées vers la zone active de la membrane présynaptique. Cette étape de mobilisation est dépendante du cytosquelette et de l’ATP. De plus, le complexe Sec6/8 (743 kDa) assurerait la spécificité d’adressage des vésicules synaptiques à la membrane présynaptique et interviendrait dans la formation des complexes SNAREs. Ce complexe coimmunoprécipite avec les septines, qui sont des composants du cytosquelette cortical et qui jouent un rôle important dans l’orientation des protéines vers la membrane plasmique 〚28〛.
L’arrimage des vésicules synaptiques à la membrane présynaptique (docking) n’a pas lieu n’importe où, mais à proximité des canaux calciques, dans la zone active (Fig. 2). De nombreuses protéines de grande taille s’assembleraient en un échafaudage permettant un amarrage fort des vésicules synaptiques. Trois protéines ont été proposées pour former cet échafaudage: aczonin (550 kDa), bassoon (550 kDa) et piccolo 〚34, 56, 58〛.
Mécanisme d’action moléculaire des neurotoxines botuliques et tétaniques. La chaîne légère des neurotoxines exerce le blocage de la libération de neuromédiateur par une activité protéolytique vis-à-vis d’une des trois protéines (synaptobrévine, SNAP25 et syntaxine) impliquées dans l’assemblage des complexes SNAREs, qui jouent un rôle essentiel dans le processus d’exocytose. La synaptobrevine ou VAMP est associée à la membrane des vésicules synaptiques, qui contiennent le neuromédiateur ; SNAP25 et syntaxine sont ancrées dans la membrane présynaptique. Cependant, des complexes SNAREs cis résultant de la phase de recyclage sont présents sur les membranes des vésicules synaptiques. Des facteurs solubles (NSF qui est une ATPase et α-SNAPs) dissocient les complexes cis et permet la formation de complexes trans entre les protéines SNAREs des membranes vésiculaires et présynaptiques. Au cours de cette phase, les protéines SNAREs, non assemblées en complexe, sont sensibles à l’activité protéasique des neurotoxines. Les complexes SNAREs permettent l’arrimage des vésicules synaptiques à la membrane présynaptique à proximité des canaux calciques. Un signal de dépolarisation entraîne une entrée de Ca++, qui est capté par la protéine calcium-senseur (synaptotagmine). Celle-ci induirait un changement de conformation des complexes SNAREs, qui déclencherait le processus de fusion des membranes vésiculaire et présynaptique, ainsi que l’exocytose, dans un temps très bref (0,3–1 ms). Pour plus de simplicité, une seule des protéines SNAREs vésiculaire ou plasmatique est représentée.
Le complexe SNARE 7 s serait en partie présent sur les membranes des vésicules synaptiques (complexe cis non fonctionnel) et préviendrait l’interaction avec la membrane présynaptique. La liaison avec NSF et αSNAPs génère la formation du complexe 20 s, qui désassemble ce complexe par l’hydrolyse de l’ATP « médiée » par NSF. Les protéines VAMP seraient alors disponibles pour former un complexe SNARE (complexe trans) avec les protéines de la membrane présynaptique (SNAP25 et syntaxine). Ce serait au cours de cette étape que les protéines SNAREs, sous forme monomérique, seraient sensibles au clivage par les neurotoxines tétanique et botulique. Le réassemblage des complexes SNAREs en complexe trans rapprocherait les vésicules synaptiques de la membrane présynaptique (docking) en les mettant sous tension, prêtes à fusionner (priming).
L’étape de priming fait également intervenir les PITPs (Phosphatidylinositol Transfer Protein), qui recrutent le phosphatidylinositol-4-phosphate (PI(4)P) pour le présenter à la phosphatidylinositol kinase (PI(4)P5K) et générer ainsi du phosphatidylinositol biphosphate (PI(4,5)P2). Ce dernier a un rôle essentiel dans les étapes de fusion membranaire. Les PITPs réguleraient également les interactions entre les protéines SNAREs 〚30〛. D’autres processus de phosphorylation ont été proposés dans l’étape ATP-dépendante du priming des vésicules synaptiques. En effet, les protéines synaptobrévine, SNAP25, syntaxine, synaptotagmine, Munc18 sont des substrats de kinases comme la protéine kinase II, dépendante de la calmoduline, la protéine kinase A et la protéine kinase C. La signification de ces phosphorylations reste à préciser 〚30〛.
Munc18 (ou nSEC1) s’associe avec une forte affinité à la syntaxine. La liaison de la syntaxine à la synaptobrevine et à SNAP25 fait intervenir le motif SNARE en C-terminal de la molécule, alors que la majeure partie cytoplasmique de la syntaxine est impliquée dans l’association avec Munc18. Munc18 se lie à la syntaxine dans sa forme fermée et prévient son association avec la synaptobrevine et SNAP25 ; inversement, lorsque la syntaxine (forme ouverte) est engagée dans le complexe SNARE, elle ne peut pas s’associer à Munc18 〚14, 36〛. En présence des vésicules synaptiques, l’action de Rab/Rab effecteur induirait un changement de conformation de Munc18 et dissocierait le complexe Munc18–syntaxine, favorisant ainsi la formation du complexe SNARE trans (VAMP, SNAP25 et syntaxine). De plus, l’analogue Munc13, qui interagit avec Munc18, pourrait convertir la forme fermée de la syntaxine liée à Munc18 en sa forme ouverte capable de former le SNARE complexe 〚30〛.
Un potentiel d’action se traduit par une entrée de Ca++ par les canaux calciques de la zone active de la membrane présynaptique, qui déclenche la fusion membranaire entre vésicules synaptiques et membrane présynaptique et la libération de neuromédiateur dans la fente synaptique dans un délai de 0,3 ms environ (Fig. 1). La synaptotagmine I, qui a la propriété de fixer le Ca++, joue un rôle essentiel dans l’étape de fusion. L’influx de Ca++ pourrait induire un changement de liaison de la synaptotagmine au complexe SNARE, ou un changement de conformation de ces protéines. La liaison aux phospholipides et à la syntaxine ainsi que la multimérisation de la synaptotagmine pourraient intervenir dans l’étape de fusion dépendante du Ca++ 〚23, 31〛. D’autres protéines pourraient avoir un rôle calcium senseur telles que DOC2 (double C2 domain), UNC13, les effecteurs de Rab3, Rabphilin et RIM (Rab Interacting Molecule), les annexines et CAPS (Calcium-dependent Activator Protein for Secretion) 〚31〛. Rab3 régulerait l’étape de fusion, en limitant le nombre de vésicules synaptiques capables de fusionner en réponse à un influx de Ca++, de façon à obtenir un signal limité dans le temps et reproductible. La forme active de Rab3 (Rab3-GTP) s’associe aux vésicules synaptiques et recrute ses effecteurs, Rabphiline et RIM. Rabphiline se lie à Rab3 sur toutes les vésicules synaptiques, alors que RIM s’associe à Rab3 uniquement sur les vésicules arrimées à la membrane présynaptique. L’activité régulatrice de ces protéines reste à définir 〚23〛.
L’étape de fusion membranaire permettant la formation d’un pore et la libération du neuromédiateur est loin d’être comprise en détail. La GTPase ARF6 interviendrait en activant la phospholipase D, qui hydrolyse la phosphatidylcholine en acide phosphatidique et en choline. L’acide phosphatidique régule la PI(4)P5K, qui synthétise le PIP2. Celui-ci module l’activité des protéines liant l’actine en région corticale et est impliqué dans le réarrangement des fibres d’actine nécessaires pour l’exocytose. Par ailleurs, une élévation locale d’acide phosphatidique en présence de Ca++ induit une courbure de la membrane lipidique et faciliterait la fusion entre membrane de la vésicule synaptique et plasmatique et, ainsi, la formation de pores 〚30, 57〛.
NSF et αSNAPS interviendraient à nouveau pour dissocier les complexes SNAREs, et recycler VAMP dans la membrane vésiculaire ainsi que SNAP25 et la syntaxine dans la membrane présynatipque. Cependant, une partie de SNAP25 et de la syntaxine resterait dans la membrane vésiculaire sous forme de complexes cis non fonctionnels. Les vésicules synaptiques sont ensuite recyclées par endocytose et rechargées en neuromédiateur 〚31, 47〛.
Lorsque VAMP, SNAP25 ou la syntaxine est clivée par une neurotoxine clostridienne, elle peut former des complexes SNAREs, mais ceux ci ne sont pas fonctionnels. La fusion des vésicules avec la membrane présynaptique n’intervient alors pas, et il n’y a pas de libération de neuromédiateur. Les protéines SNAREs jouent un rôle essentiel dans la fusion membranaire permettant la neuroexocytose.
A9 Particularités du blocage de la neuroexocytose par les différentes neurotoxines clostridiennes
Les neurotoxines botuliques et tétanique sont les toxines les plus puissantes connues. La dose mortelle de neurotoxine botulique A chez l’homme adulte est estimée entre 0,1 et 1 μg par voie orale 〚46〛, et à 10 doses létales100 souris (soit 250 ± 150 pg) pour la neurotoxine tétanique à la suite d’un tétanos survenu après blessure 〚44〛. Pourtant, leur activité enzymatique n’est pas très élevée. Le Km de la neurotoxine tétanique pour son substrat VAMP est d’environ 200 μM. Or, environ six molécules de neurotoxine tétanique par micromètre cube dans le neurone sont suffisantes pour provoquer un blocage total de la libération de neurotransmetteur en 2–3 h, alors que le nombre de molécules de VAMP est estimé entre 20 000 et 50 000 par micromètre cube 〚43〛. En fait, VAMP, comme les autres partenaires SNAP25 et syntaxine, ne se dissocie du complexe SNARE que pendant un court instant (1 à 5 s), lors de l’arrimage des vésicules sur la membrane présynaptique ; ce n’est qu’au cours de cette phase qu’elles sont accessibles aux neurotoxines clostridiennes. Comme ces protéines sont recyclées, les neurotoxines clivent progressivement la totalité des molécules substrats. Mais les signes cliniques apparaissent bien avant. Chez l’animal, une inhibition de libération d’acétylcholine de 20 % au niveau de la jonction neuromusculaire est suffisante pour prévenir la contraction musculaire. De plus, quand 10–15 % des fibres musculaires du diaphragme sont paralysées, des troubles respiratoires, avec asphyxie, surviennent 〚43〛. Cela rend compte de l’extrême potentialité des neurotoxines clostridiennes chez l’animal.
Les neurotoxines clostridiennes n’altèrent pas la structure des synapses. La seule modification morphologique consiste en une augmentation du nombre de vésicules synaptiques à proximité de la membrane présynaptique. D’un point de vue électrophysiologique, les neurotoxines clostridiennes bloquent la libération stimulée de neuromédiateur. Dans les conditions physiologiques, cette libération stimulée est synchrone, c’est-à-dire qu’un grand nombre de vésicules synaptiques libèrent leur contenu en neuromédiateur en même temps en réponse à une stimulation (entrée de Ca++ à l’extrémité nerveuse). Lors de l’intoxication par les neurotoxines botuliques A, E et C, dont la cible est SNAP25, et aussi par la syntaxine pour le type C, la libération d’acétylcholine reste synchrone. Mais les neurotoxines botuliques B, D et F ainsi que la neurotoxine tétanique, dont le substrat est VAMP, provoquent une désynchronisation importante 〚47〛.
Les aminopyridines sont des agents bloquant des canaux potassiques. Elles provoquent indirectement une entrée de Ca++ et, par conséquent, augmentent la libération stimulée de neuromédiateur. Ces agents synchronisent et restaurent partiellement la neurotransmission des jonctions neuromusculaires intoxiquées par les neurotoxines botuliques A, E et C, alors qu’ils sont inefficaces dans l’intoxication par les neurotoxines botuliques B, D et F ainsi que par la neurotoxine tétanique.
Les neurotoxines botuliques A et E clivent le même substrat SNAP25, mais sur deux sites différents. La neurotoxine botulique A relargue neuf acides aminés C-terminaux, tandis que la neurotoxine botulique E excise un plus long peptide (25 résidus). Les effets cliniques de ces deux toxines sont différents. La neurotoxine botulique A induit une paralysie de longue durée (plusieurs mois), alors que l’intoxication par la neurotoxine botulique E est courte (quelques semaines) 〚15〛. Ceci pourrait être dû à une plus longue persistance de la neurotoxine botulique A dans les jonctions neuromusculaires par rapport à la neurotoxine botulique E. Cependant, la durée de vie des deux toxines semble identique. Dans les cellules chromaffines, il a été montré que la neurotoxine botulique A peut persister au moins pendant 21 jours, alors que la persistance de la neurotoxine botulique B et de la neurotoxine tétanique est plus courte 〚39〛. La différence de durée d’intoxication est fonction de la rapidité de remplacement de SNAP25, modifié par les neurotoxines dans les jonctions neuromusculaires. SNAP25, tronquée par la neurotoxine botulique A, persiste pendant une plus longue période, alors que SNAP25, clivée par le type E, est remplacée rapidement par la forme native fonctionnelle. La modification par la neurotoxine botulique A altère peu SNAP25, qui n’est reconnu que tardivement par le système de réparation de la cellule, mais cette altération est suffisante pour inhiber sa fonction. Au contraire, la protéolyse plus large par la neurotoxine botulique E induit une rapide néosynthèse de SNAP25 〚15, 37〛.
Au cours de l’intoxication par la neurotoxine botulique A, des bourgeons secondaires se développent aux extrémités nerveuses intoxiquées, au niveau des boutons synaptiques et des nœuds de Ranvier. Le muscle paralysé s’atrophie transitoirement et secrète des facteurs trophiques, qui sont probablement responsables du bourgeonnement. Les bourgeons nerveux reforment des jonctions neuromusculaires fonctionnelles. Plus tard, la jonction neuromusculaire initiale retrouve une morphologie et une activité normales, et les bourgeons secondaires dégénèrent 〚9〛.