

Abridged English version
Pleistocene climatic oscillations are known to be an important motor in European terrestrial fauna and flora. During cold episodes, several refuges have functioned in the southern peninsulas (Iberian and Italian peninsulas, Balkans), but also further east on the foothills of Caucasus. Recently, such refuges have been shown to produce two different subspecies in the eastern Mediterranean short-tailed mouse, yielding the two subspecies M. macedonicus macedonicus and M. m. spretoides. In the Mediterranean island of Cyprus, it was known that two species of mice co-existed, the House mouse M. musculus domesticus and another form until now referred to as M. macedonicus from superficial morphological inspection. Recently, a morphological analysis of recent and subfossil specimens showed that the animals referable to this second taxon were indeed clearly distinguishable from M. macedonicus. Here, we characterise genetically seven mouse specimens from Cyprus using 310 pb of mitochondrial 16s-rDNA, 230 pb of the variable part of the mitochondrial D-loop, and 761 pb of the Abpa nuclear gene 2nd intron. We compare the sequences obtained with that of the mound builder mouse M. spicilegus, the close relative species of M. macedonicus. The molecular divergence obtained and the phylogenetic trees resulting from this comparison clearly show that five Cyprus animals are almost equidistant from M. macedonicus than it is from M. spicilegus, both for mitochondrial and nuclear genes (the levels of mitochondrial nucleotidic divergence observed – about 8% for the D-Loop and 5% for the 16S rRNA – correspond to at least several hundred thousand years). The divergences observed at the Abpa 2nd intron were ten times smaller (0.6%).
Altogether, these results clearly indicate that these individuals belong to a new taxon, differentiated from the other two short-tailed species.
This new species, designated here provisionally as M. sp. nova, deserves further description of type specimens before it is assigned a name, which possibly could be M. cypriacus. Indeed, two hypotheses can be formulated as to its origin.
- (i) It is a true endemic having reached Cyprus long before the last glacial episode. This could be in keeping with the observation of fossil mice in mid-Pleistocene deposits and would imply a colonisation of the island by some rare accidental rafting event. Indeed, the island is separated from the mainland by a deep channel that was never emerged, even during the glacial-period drop-off.
- (ii) It is a quite recent invader that colonised the island with its first human settlers along with the commensal M. m. domesticus. In this case, its origin would have to be found in the nearby mainland, where a localised refuge different from the two others, responsible for the existence of the two subspecies of Mus macedonicus, could have functioned on the southern coast of Asia Minor.
Our results point out toward the complex functioning of the faunistic refuges having existed in the Near East, a point also underlined by recent phylogeographic data on other terrestrial species such as the European hedgehog.
1 Introduction
Les oscillations climatiques liées aux périodes glaciaires ont engendré des mouvements concomitants des faunes terrestres très importants. Dans le domaine paléarctique continental européen, notamment, ces mouvements se sont traduits par le fonctionnement de nombreux refuges dans les péninsules du Sud de l'Europe (Ibérique, Italique, Balkanique) [1], mais il a été également montré que de tels refuges avaient fonctionné plus à l'est, en Asie Mineure [2–4]. Un des apports majeurs de la phylogéographie moléculaire a été de pouvoir étudier les conséquences génétiques de ces alternances de phases d'isolement et d'expansion, et de mettre clairement en évidence leur rôle dans la création d'espèces vicariantes, puis sympatriques. Chez les souris du genre Mus, il a été récemment proposé [3] que le fonctionnement de refuges distincts situés au sud du Caucase et autour de la mer Noire était responsable de la différentiation génétique des espèces Mus spicilegus et Mus macedonicus, ainsi que de l'existence de deux sous-espèces Mus macedonicus macedonicus et Mus macedonicus spretoides, occupant respectivement le Nord et le Sud de l'aire de répartition de ce taxon.
D'après la littérature, Chypre, grande ı̂le océanique de Méditerranée, se situe théoriquement dans l'aire de répartition de Mus macedonicus [5,6] où celle-ci vivrait en sympatrie avec la sous-espèce commensale Mus musculus domesticus. Une analyse morphométrique récente, portant sur la distinction interspécifique d'un assemblage de muridés issu d'un site Néolithique précéramique (fin du 9e millénaire av. J.-C.), a en effet démontré que deux espèces sympatriques occupaient déjà l'ı̂le à cette période [7]. La première appartient clairement à la sous-espèce commensale. La seconde présente une distance morphologique significative avec les souris sauvages actuelles d'Israël (Mus macedonicus spretoides), qui avaient servi de référence pour le Proche-Orient [7]. Ceci fut interprété selon trois hypothèses :
- (i) la survivance d'une des deux formes du genre Mus décrites par Bate [8] et Boeschoten et Sondaar [9] parmi la faune Pléistocène découverte dans cinq grottes ossifères dans la région de Cape Pyla ;
- (ii) une origine géographique différente de celle des macedonicus d'Israël, seule sous-espèce avec laquelle la forme chypriote avait été comparée ;
- (iii) la conséquence d'une dérive morphologique holocène liée aux effets stochastiques d'une fondation à partir de quelques individus fortuitement immigrés.
Il était donc intéressant de comprendre les relations de parentés entre ces différents taxons et d'aborder la question de l'origine du peuplement de l'espèce non commensale à Chypre. Dans la présente étude, nous analysons génétiquement six individus actuels en provenance de cette ı̂le et montrons que cinq d'entre eux appartiennent à une espèce encore jamais décrite. Nous discutons ces résultats à la lumière du fonctionnement possible des refuges glaciaires ayant existé en Asie Mineure.
2 Matériel et méthodes
Onze individus, dont six en provenance de Chypre, ont été analysés (Tableau 1).
Origine géographique des échantillons analysés
| Nom des taxons sur la Fig. 1A | Pays | Localités | Espèces |
| Chypre 1 | Chypre | Kissonerga | Mus sp. nova |
| Chypre 2 | Chypre | Mandria | Mus sp. nova |
| Chypre 3 | Chypre | Trombovounos | Mus sp. nova |
| Chypre 4 | Chypre | Ayios Dometios | Mus sp. nova |
| Chypre 5 | Chypre | Kophinou | Mus sp. nova |
| dom-Chypre 6 | Chypre | Kissonerga | Mus musculus domesticus |
| dom-France | France | Les Embiez | Mus musculus domesticus |
| musculus | Bulgarie | Sokolovo | Mus musculus musculus |
| macedonicus | Bulgarie | Jaboltchevo | Mus macedonicus macedonicus |
| spretoı̈des | Israël | Mont Carmel | Mus macedonicus spretoides |
| spicilegus | Serbie | Pančevo | Mus spicilegus |
| Nom des taxons sur la Fig. 1B | Pays | Localités | Espèces |
| Chypre 3 | Chypre | Trombovounos | Mus sp. nova |
| dom-Chypre 6 | Chypre | Kissonerga | Mus musculus domesticus |
| dom-France | France | Les Embiez | Mus musculus domesticus |
| musculus | Pologne | Bialowieza | Mus musculus musculus |
| macedonicus | Bulgarie | Slantchev Briag | Mus macedonicus macedonicus |
| spretoı̈des | Israël | Mont Carmel | Mus macedonicus spretoides |
| spicilegus | Serbie | Pančevo | Mus spicilegus |
L'ADN a été extrait à partir de mandibules récoltées dans les pelotes de réjection de chouette effraie (Tyto alba) ou à partir d'un fragment de rate avec le kit DNeasy Tissue QIAGEN. Onze échantillons ont été typés pour l'ADN mitochondrial, et sept pour l'ADN nucléaire.
2.1 ADN mitochondrial
270 pb du gène codant pour l'ARNr 16S ont été amplifiées dans la région la plus variable à partir des amorces précédemment définies [10] entre les positions 1057 et 1329 de la séquence référence de Mus domesticus [11], ainsi que 310 pb de la région hypervariable de la D-loop avec ce couple d'amorces : MTL3L (5′-CACCACCAGCACCCAAAGCT-3′) et MTL4R (5′-TTGGTTTCACGGAGGATGGT-3′). Les produits de PCR ont été séquencés sur séquenceur automatique ALF AP Biotech. Les séquences correspondantes ont été concaténées et les analyses effectuées sur un total de 494 sites (numéros d'accession dans GenBank AY562503 à AY562513 pour la D-loop et AY562521 à AY562531 pour l'ARNr 16S).
2.2 Gène nucléaire
Un fragment de 1235 pb du gène Abpa a été amplifié avec les amorces ABPA1L (5′-AGCTCACTGGTGCTCTCTTGC-3′) et ABPA1R (5′-CTTAGACACCTGGGGAGAAGG-3′). L'exon 2 (188 pb) ainsi que l'intron 2 (761 pb) ont été séquencés [12]. Les numéros d'accession dans GenBank sont AY562514 à AY562520.
La phylogénie de ces haplotypes a été réalisée par la méthode du neighbour-joining [13] appliquée à une matrice de divergence nucléotidique corrigée (Kimura 2-paramètres) grâce au logiciel Phylo-Win [14]. La robustesse des arbres a été évaluée par bootstrap (1000 réplications).
3 Résultats
Nous présentons sur la Fig. 1A l'arbre phylogénétique représentant la différenciation mitochondriale des échantillons chypriotes comparée à celles de plusieurs autres taxons proches. Il apparaı̂t clairement que le groupe des individus chypriotes de la forme non commensale se situe à l'intérieur du groupe spicilegus/macedonicus, tout en étant à peu près aussi divergent de l'un et l'autre taxon (les valeurs de divergence nucléotidique moyennes pour le fragment de D-loop s'établissent à 0,073, 0,083 et 0,097 pour spicilegus/macedonicus, macedonicus/Chypre, et spicilegus/Chypre respectivement). Pour le fragment d'ADNr-16S, nous obtenons 0,052, 0,047 et 0,039.
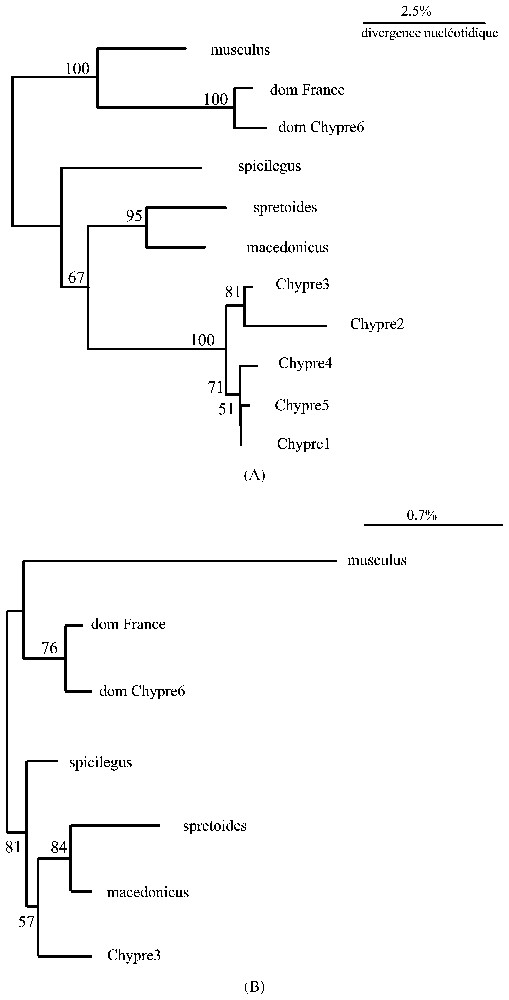
(A) Phylogénie mitochondriale (ADNr16S & D-Loop) des six échantillons de Chypre et des autres taxons proches. (B) Phylogénie générée à partir des 661 pb de l'intron n∘ 2 du gène nucléaire Abpa. La phylogénie de ces haplotypes a été réalisée par la méthode du neighbour-joining appliquée à une matrice de divergence nucléotidique corrigée (Kimura 2-paramètres). La robustesse des nœuds a été évaluée par bootstrap (1000 réplications).
Dans la Fig. 1B, nous montrons l'arbre obtenu pour les 661 pb de l'intron 2 du gène nucléaire Abpa. Bien que les divergences nucléotidiques soient modérées (les valeurs de divergence nucléotidique moyenne s'établissent à 0,007, 0,007 et 0,005 pour spicilegus/macedonicus, macedonicus/Chypre, et spicilegus/Chypre respectivement), elles montrent clairement que chaque taxon possède une séquence différente, et l'arbre produit a exactement la même topologie que celui de la Fig. 1A.
L'ensemble de ces résultats montre que deux espèces distinctes sont bien présentes en sympatrie sur l'ı̂le : la souris domestique M. musculus domesticus d'une part, et d'autre part un nouveau taxon clairement apparenté à M. spicilegus et M. macedonicus, mais aussi différencié de ces deux derniers qu'ils le sont l'un de l'autre. Ces résultats suggèrent donc fortement que ce taxon est une espèce à part entière, qui ne correspond ni à M. spicilegus ni à M. macedonicus. La question de l'origine, insulaire ou non, de cette espèce reste posée.
4 Discussion
4.1 Divergence moléculaire
Les calibrations temporelles disponibles dans la littérature pour les régions étudiées de l'ADN mitochondrial ne nous permettent pas de faire une estimation précise des temps de divergence séparant les trois taxons spicilegus/macedonicus/sp. nova. Néanmoins, si l'on considère que la nouvelle espèce se situe à équidistance des deux autres, on peut se référer au chiffre d'environ 1,1 Ma proposé par She et al. [15] sur la base d'une étude d'hybridation ADN/ADN, calibrée par une divergence Mus/Rattus située aux alentours de 10 Ma. Quelle que soit la vraie valeur de ce chiffre, elle est synonyme en tout cas d'une séparation des taxons largement antérieure à la dernière glaciation (50 000 15 000 AC).
Pour l'intron 2 du gène de la protéine ABP, les divergences moléculaires sont très faibles, mais du même ordre entre les trois taxons (div. moyenne de 0,8 %, ce qui est dix fois plus faible que les valeurs observées pour la DLoop). Notons que la partie exonique que nous avons séquencée (188 pb de l'exon 2, non montré) ne présentait aucune divergence entre les trois espèces, contrairement à ce qui a été observé chez M. spretus et M. musculus [12].
4.2 Origine du taxon chypriote
Nos données montrent donc clairement l'existence d'un troisième taxon grosso modo équidivergent de l'une et l'autre espèces jumelles M. macedonicus et M. spicilegus. A priori, plusieurs scénarios sont possibles pour son origine :
- (i) Il s'agit d'une espèce véritablement endémique depuis longtemps à Chypre et qui aurait survécu jusqu'à nos jours. Elle serait donc, avec Crocidura zimmermanni (Crète, [16]), la seule espèce mammalienne endémique pléistocène des ı̂les méditerranéennes à avoir subsisté. Elle aurait colonisé l'ı̂le à la faveur d'un événement rare de transport par radeau au cours du Quaternaire. D'après les données géologiques et paléontologiques, il n'a jamais existé de connexion terrestre entre l'ı̂le et le continent depuis le Miocène [17], et l'absence de fossiles du Tertiaire suggère que l'ı̂le était déjà séparée du continent lors de l'assèchement messinien (–6 Ma) [18,19]. Bien que la recherche portant sur les fluctuations fines des niveaux marins du Pléistocène n'en soit qu'à ses débuts, il ne fait pas de doute que Chypre était séparée par de larges chenaux d'eau profonde lors des régressions marines associées au dernier maximum glaciaire, comme elle l'est encore aujourd'hui [20]. L'endémisme de cette espèce pourrait être corroboré par l'existence des formes du genre Mus décrites parmi la faune des gisements pléistocènes de Chypre [9] ainsi que par la mention d'une forme sauvage dans un niveau d'occupation épipaléolithique (10 500 BC) de l'abri sous roche d'Akrotiri-Aetokremnos [21]. L'analyse morphologique de ces fossiles est en cours (Cucchi, obs.pers).
- (ii) L'espèce trouve son origine ailleurs sur le proche continent, où aurait fonctionné un refuge l'isolant de M. macedonicus. Elle aurait alors colonisé l'ı̂le à la faveur des premiers mouvements humains, en même temps ou peu avant M. m. domesticus. Cette occurrence poserait alors la question de la possibilité d'un transport passif et occasionnel d'espèces non strictement commensales lors des déplacements exploratoires de petits groupes humains épipaléolithiques. Chypre et les autres ı̂les méditerannennes ont été définitivement colonisées au Néolithique.
Les données disponibles ne permettent pas de trancher pour le moment entre les deux scénarios. L'analyse morphologique des fossiles pléistocènes de Chypre ainsi que l'exploration des isolats géographiques d'Asie Mineure devraient permettre de lever le voile. Cette nouvelle espèce, qui doit faire l'objet d'une description morphologique détaillée de spécimens types, pourrait alors être nommée Mus cypriacus.