

1 Introduction
Because biodiversity conservation and management are worldwide concerns, it is essential to possess reliable, replicable, and inexpensive means of monitoring different target organisms. Due to their abundance, diversity and sensitivity to environmental changes (such as habitat fragmentation), ants are good bio-indicators. They are also considered to be a keystone taxon in the functioning of terrestrial ecosystems due to their impact on such things as soil ventilation, draining and renewal, nutrient recycling, predation, plant protection, and seed dispersal [1,2].
Given ants' high species richness and ubiquity, many techniques have been developed to collect them, but these sampling methods have mostly been used in the tropics. They include the use of (1) a variety of types of extractors for soil and/or leaf-litter (e.g., Berlese–Tullgren or Winkler extractors), (2) pitfall traps to collect ants on the ground, (3) baits to attract different varieties of foraging guilds, (4) canopy fogging, (5) vegetation beating, and (6) manual collection [3–5]. While each of these methods on its own can provide different types of bio-ecological information on species [6], combining them permits researchers to obtain the most complete information possible on population densities and species richness [7–9]. Nevertheless, each method and/or any combination of them can also influence the nature of the results so that their usefulness depends on the questions we are trying to answer; for example, in tropical environments, manual collection (or direct sampling), the use of baits, pitfall traps (for open microhabitats), plus the Winkler extractor (for litter-dwelling ants) form a good combination of methods for assessing biodiversity by guaranteeing a thorough representation of the ant fauna and the comparability of samples [7]. Delabie et al. [10] compared the efficiency of 17 different ant-sampling methods in the Brazilian Atlantic forest, and showed that the combination of pitfall traps and leaf-litter sampling with the Winkler extractor permitted 50% of the total known species to be captured.
The objectives of this study were twofold. First, we conducted sampling in the ‘Parc naturel régional des Grands Causses’, a part of an area in southern France that, in association with the ‘Parc national des Cévennes’, is a candidate to become a UNESCO ‘world heritage’ site. We therefore compared three vegetal formations representative of the area: a steppic lawn (‘causse’, sensu stricto), an oak grove, and a pine forest. Second, because most studies on ant diversity and distribution are conducted in the tropics, we questioned if the methods used in that context could be transferred to temperate regions that have received little attention until now [11]. We therefore aimed to compare the efficiency of four sampling methods (i.e., the Winkler extractor, pitfall traps, baiting and manual collection) and to determine the optimal combination of ant-sampling techniques.
2 Material and methods
2.1 Study sites
The Grands Causses (average altitude 800 m) are located south of the Massif Central (southern France) and include the Causse de Méjean, the Causse noir, the Larzac, and the Causse de Sauveterre. Only the latter three are part of the ‘Parc naturel régional des Grands Causses’, the Causse de Méjean being a part of the ‘Parc national des Cévennes’ [12,13]. The Grands Causses, sparsely populated, are wide, dry calcareous table lands cut by deep canyons (karstic system) characterized by harsh winters and subject to low temperatures and dry summers with strong thermal amplitude and a vegetation adapted to drought. They contain vast settlements of steppic lawns (causse) and woodland pines, in particular in their eastern and septentrional zones, while pubescent oak spontaneously colonizes the most exposed slopes.
Ant sampling was carried out in May 2005 around the village of Saint-Léons-en-Levezou (department of the Aveyron) by 35 people (the authors plus graduate students from the University of Toulouse-3). We sampled a causse (centre of the parcel: , ; altitude: 850 m) characterized by shrub and herbaceous plant species, such as Amelanchier ovalis, Bromus erectus, Buxus sempervirens, and Rubus fructicosus, an oak grove (, ; altitude: 770 m) dominated by Quercus pubescens, and a pine forest of Pinus sylvestris (, ; altitude: 780 m). The distance between (1) the causse and the oak grove was 2.2 km, (2) between the causse and the pine forest, 4.5 km, and (3) between the oak grove and the pine forest, 4.3 km. The leaf-litter layer was thick in the oak grove and pine forest (up to 12 cm), but much more limited in the causse (up to 3 cm).
2.2 Sampling methods
Based on recommendations presented in previous studies in the tropics [7,14], we used four sampling methods: Winkler extractors, pitfall traps, baiting and manual collection. Sampling was conducted in the centre of each of the three distinct vegetal formations on six adjacent transects 125 m long and 25 m wide (1.875 ha), each divided into five 25 m ×25 m parcels (30 parcels in total) where the four sampling methods were used. All methods were applied randomly in each parcel, leading to 30 replicates for each environment. The total time spent sampling with all methods was about 400 h (all-pooled).
For manual collection, four observers walked randomly around each parcel during 20 min (1 h 40 min of sampling effort per transect or 10 h per vegetal formation) and collected ants foraging on trees or on the ground, plus colonies nesting in dead wood or under rocks. Baits (one piece about 2 cm3 of tinned sardines; 30 baits per parcel) were placed on absorbent paper () on the ground for 3 h [3]. Pitfall traps (30 per parcel) consisted of plastic 75-mm-diameter cups (a size recommended by Bestelmeyer et al. (2000) [7] to optimize trap efficiency) filled to one-third of their height with slightly (5%) soapy water (neither attractive nor repulsive to ants). They were placed randomly, but more than 1 m from the edges of each parcel to avoid interference between parcels, and the vegetation around them was not cleared. The traps were removed 24 h later and the ants collected. Finally, we sifted the litter from a 1-m2 quadrate in each parcel, and placed it in a numbered bag. In the laboratory, the sifted litter was placed into the inner mesh bag of a Winkler extractor. This bag was then introduced into the outer, funnel-shaped canvas bag whose end is connected to a cup containing 80° ethanol. The extractor was then hung in a well-aired environment, out of direct sunlight. In response to the increasing levels of dryness, the arthropods, including ants, crawl out of the inner mesh bag, and then fall into the ethanol [7]. All the resulting samples were first coded and then put into small plastic tubes filled with 80° ethanol. Later the ants were mounted in order to constitute a reference collection and identified according to Bolton [15,16]). Voucher specimens were deposited at Micropolis (Saint-Léons-en-Levezou, France), the ‘Laboratoire d’évolution et diversité biologique', University Toulouse-3 (France), and the ‘Laboratório de Mirmecologia’, Cocoa Research Center CEPEC/CEPLAC (Ilhéus, Bahia, Brazil).
2.3 Statistical analyses
Species richness (number of collected species) and species occurrences (number of times that a given species was collected in all samples using the same method) were compared between vegetal formations and sampling methods. To estimate and compare the efficiency of the sampling methods, rarefaction curves (cumulated numbers of species according to the number of occurrences for each species) were calculated for each vegetal formation, each method, and all methods combined. Because comparing sampling techniques on the basis of number of samples has little meaning (no two methods having the same probability of capture) and because the number of individuals is not a reliable parameter in ant studies (data can be distorted when ants are collected close to a nest, an ant trail or a site where ants recruit nest mates), we took into account only species occurrences. Matrices of species occurrences (presence/absence) according to the area where the ants were sampled and to sampling methods were treated using the EstimateS 7.1 software [17] with 500 randomizations of the sampling order without replacement (see [18]).
To sort the samples (i.e. type of environment and sampling methods) according to the ant species they host (see Appendix A), we used the Self-Organizing Map algorithm (SOM Toolbox version 2 for Matlab®; http://www.cis.hut.fi/projects/somtoolbox/documentation/; see [19,20]). Combining clustering and functions, the SOM is a powerful tool used to visualize high-dimensional ecological data; more specifically, this technique is particularly relevant to patterning species assemblages using data on species occurrences. The SOM algorithm is an unsupervised learning procedure that transforms the multidimensional input data constituted here by 33 nodes (one per ant species) connected to the 273 samples into a two-dimensional map (shown as 80 hexagonal cells) subject to a topological constraint. The SOM plots the data so that similar samples are found together on the grid and, inversely, very different samples are far from each other. In this study, we used the ordination process detailed in [21] where the training was broken down into an ordering phase (the first 3000 iterations), and a fine-tuning phase (7000 iterations). At the end of the training, a species assemblage is known for each virtual site and the ‘best-matching unit’ (BMU) is determined for each sample that is set in the corresponding hexagon of the final SOM. To assess whether the map has been properly trained, the topographical error was used as a measure of map quality [20]. The intensity of the connection between input and output layers calculated during the learning process can be considered as the probability of occurrence of each ant species in the area concerned. It is represented on the SOM using a shaded scale, allowing us to analyse the effect of each variable (ant species) on the patterning input dataset (environments and sampling methods). At the end of the learning process, in order to detect the cluster boundaries on the map, the unified-matrix (U-matrix) approach [22] was applied. The U-matrix displays the distances between the virtual sites and provides a landscape formed with light plains separated by dark ravines. The sites in the plains are similar (for ant species assemblage) and clusters become clear.
Variations in the environment (e.g., vegetal formations) or the sampling method frequencies among the six clusters resulting from the SOM were analyzed using a generalized linear model [23] with a binomial error (proportion data, statistics) by using sample frequencies in each cluster as denominator vector (e.g., reference). First, models with the six clusters were fitted to the data (e.g., complete models). Second, simplified models (related to the environment or the sampling methods) grouping certain clusters were adjusted to the data and only those which were statistically similar to the complete model ( test) were retained (e.g., simplified models).
3 Results
3.1 Sampling efficiency of the ant fauna in the three vegetal formations studied
All vegetal formations and methods pooled, we recorded 33 ant species representing 13 genera and three subfamilies: Dolichoderinae (Tapinoma), Formicinae (Camponotus, Formica, Lasius and Plagiolepis), and Myrmicinae (Aphaenogaster, Temnothorax, Messor, Myrmecina, Myrmica, Solenopsis, Stenamma, and Tetramorium) (Appendix A, Table 2). We noted 29 species in the causse, 22 in the oak grove, and 17 in the pine forest. All three vegetal formations were suitably sampled because the height of the curves tends to decrease (i.e., the number of new species falls as the number of samples increases; Fig. 1). Thus, for an equal sampling effort using identical sampling techniques, each vegetal formation seems to have been globally sampled with the same effectiveness.
Number of occurrences for the ant species collected from the three environments sampled (causse, oak grove, pine forest) using the four sampling methods. Subfamilies: M=Myrmicinae; F=Formicinae; D=Dolichoderinae
| Ant species | Occurrences of species | |||
| Causse | Oak grove | Pine forest | ||
| M | Aphaenogaster gibbosa (Latreille) | <5 | <10 | 0 |
| M | Aphaenogaster subterranea (Latreille) | 0 | <10 | 0 |
| F | Camponotus aethiops (Latreille) | <10 | <10 | 0 |
| F | Camponotus ligniperdus (Latreille) | <10 | <10 | <10 |
| F | Camponotus piceus (Leach) | <10 | <10 | 0 |
| F | Formica cunicularia Latreille | 12 | 0 | 0 |
| F | Formica fusca Linnaeus | <10 | 0 | <10 |
| F | Formica gagates Latreille | <10 | 26 | <10 |
| F | Formica pratensis Retzius | <10 | <10 | 0 |
| F | Formica rufa Linnaeus | <10 | 0 | <10 |
| F | Formica sanguinea Latreille | <10 | 0 | 0 |
| F | Lasius alienus (Foerster) | <10 | <10 | <10 |
| F | Lasius brunneus (Latreille) | 0 | <10 | <10 |
| F | Lasius emarginatus (Olivier) | <10 | 0 | 0 |
| F | Lasius flavus (Fabricius) | 0 | 0 | <10 |
| F | Lasius fuliginosus (Latreille) | <10 | 13 | <10 |
| M | Temnothorax luteus Forel | <10 | <10 | 16 |
| M | Temnothorax niger Forel | <10 | 0 | 0 |
| M | Temnothorax nigriceps Mayr | <10 | <10 | 0 |
| M | Temnothorax racovitzai Bondroit | <10 | 31 | 21 |
| M | Temnothorax tuberum (Fabricius) | <10 | 0 | 0 |
| M | Messor structor (Latreille) | <10 | 0 | 0 |
| M | Myrmecina graminicola (Latreille) | <10 | <10 | 12 |
| M | Myrmica lobicornis Nylander | <10 | 0 | 0 |
| M | Myrmica ruginodis Nylander | <10 | 30 | 20 |
| M | Myrmica sabuleti Meinert | 17 | <10 | <10 |
| M | Myrmica schencki Viereck | <10 | <10 | <10 |
| F | Plagiolepis pygmaea (Latreille) | <10 | 0 | 0 |
| M | Solenopsis banyulensis Bernard | <10 | <10 | 0 |
| M | Stenamma petiolatum Emery | 0 | <10 | 0 |
| D | Tapinoma erraticum (Latreille) | <10 | <10 | <10 |
| D | Tapinoma nigerrimum (Nylander) | 23 | <10 | 11 |
| M | Tetramorium caespitum (Linnaeus) | 47 | <10 | <10 |
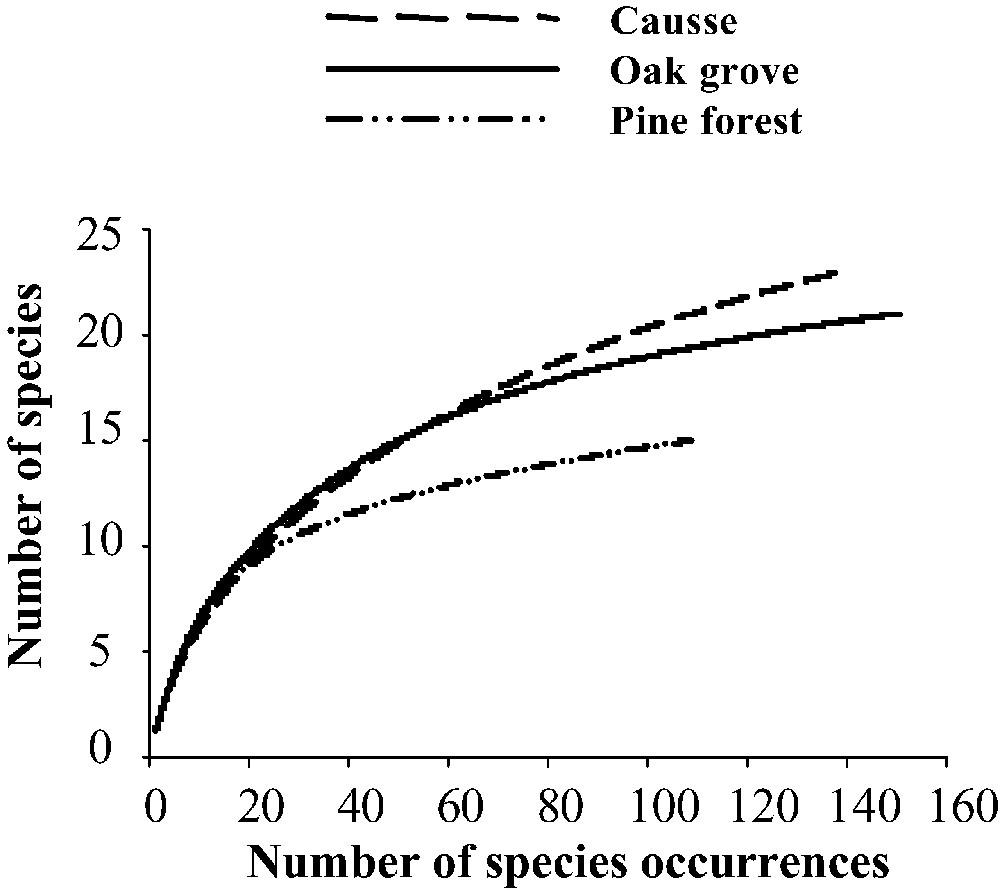
All sampling methods pooled, sample-based rarefaction curve representing the number of species occurrences for each environment sampled (causse, oak grove, pine forest) (N=90 samples for each environment). Long dashed line=causse; solid line=oak grove; short dashed line=pine forest. Curves are plotted from the means of 500 randomizations of a sample accumulation order.
3.2 Comparison of the efficiency of the four sampling methods used and optimal combination of sampling methods according to vegetal formation
Each sampling technique enabled us to collect 20 species or more (manual collection: 27; baits: 20; pitfall traps: 21; Winkler method: 25). If we compare the passive sampling methods (manual collection was not taken into account in the comparison because it is an active method), we note that none of them allowed us to collect all of the species, regardless of the study area. Because the curves are not stabilized, each technique resulted in the undersampling of the three vegetal formations, indicating that a greater sampling effort would have contributed to the addition of new species. The Winkler extractor best sampled the three vegetal formations, particularly in terms of species occurrences (Fig. 2), but was less efficacious on the causse, where there is little leaf-litter.
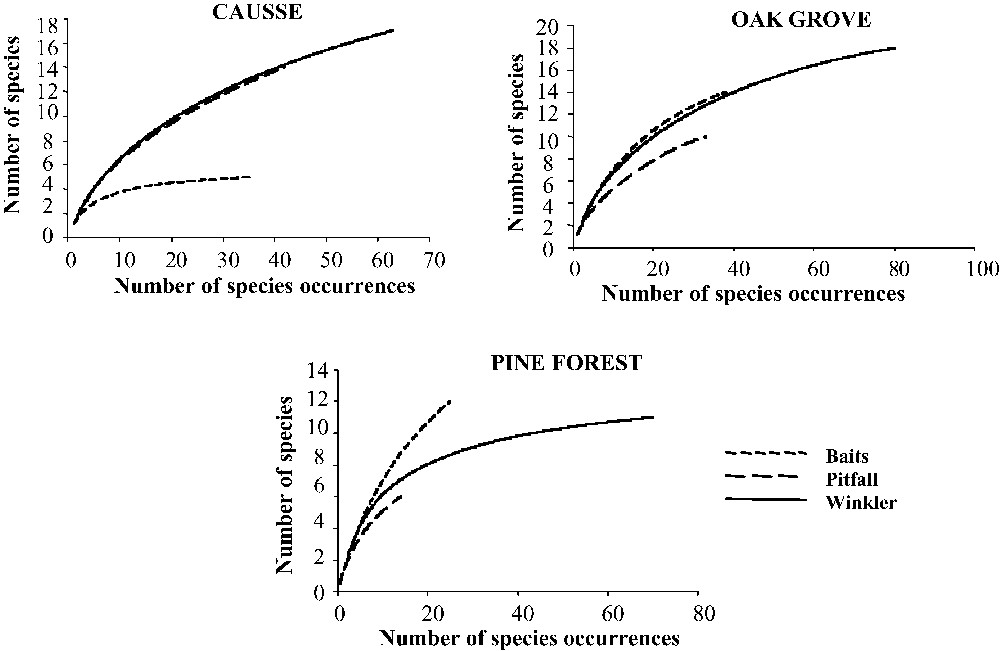
For the three passive sampling methods used (Winkler extractors, pitfall traps, baits), sample-based rarefaction curves representing the number of species occurrences from each environment (causse, oak grove, pine forest) (N=90 samples for each method used in each environment). Short dashed line=baits; long dashed line=pitfall traps; solid line=Winkler extractors. Curves are plotted from the means of 500 randomizations of a sample accumulation order.
We tried to determine the optimal combination to best sample each vegetal formation by combining two, three or four sampling methods (Table 1). For the causse, the combination of only three methods (i.e., manual collection, Winkler extractors, and pitfall traps) allowed all of the ant species to be captured, highlighting the fact that baiting did not add anything in this case. For the oak grove and the pine forest, only the combination of the four methods allowed all ant species present to be captured. Consequently, overall the four methods are complementary, each contributing a component to the total data.
Species number and percentage of the total species number based on the combination of sampling methods
| Environment | Combinations of sampling methods | Number of species | Percentages / total number of species |
| Causse | Winkler – Manual collection | 25 | 86.2 |
| Pitfall – Manual collection | 25 | 86.2 | |
| Winkler – Pitfall | 23 | 79.3 | |
| Winkler – Pitfall – Manual collection | 29 | 100.0 | |
| Oak grove | Winkler – Baits | 19 | 86.4 |
| Winkler – Pitfall | 20 | 90.9 | |
| Pitfall – Baits | 16 | 72.7 | |
| Winkler – Pitfall – Baits | 21 | 95.5 | |
| Winkler – Pitfall – Baits – Manual collection | 22 | 100.0 | |
| Pine forest | Winkler – Manual collection | 16 | 94.1 |
| Baits – Manual collection | 15 | 88.2 | |
| Winkler – Baits | 14 | 82.4 | |
| Winkler – Baits – Manual collection | 17 | 100.0 |
3.3 Influence of the ‘environment’ and ‘sampling methods’ on the sampling of ant fauna
The 80 units the SOM trained with species presence/absence data had a topographical error of 0.0001. This map thus preserved the typology of the input data well, and was relevant for subsequent interpretation (clusters A–F, Fig. 3a and b). After training the SOM with species data, the U-matrix helped to tease out six clear clusters. Thus, samples were classified into six subsets according to ant assemblages; i.e. according to the different distribution patterns, which were characteristic of each species. To reveal the relationships between environments, sampling techniques, and ants, we subsequently introduced the environmental types and sampling techniques into the SOM previously trained with the 33 ant species (Fig. 4).
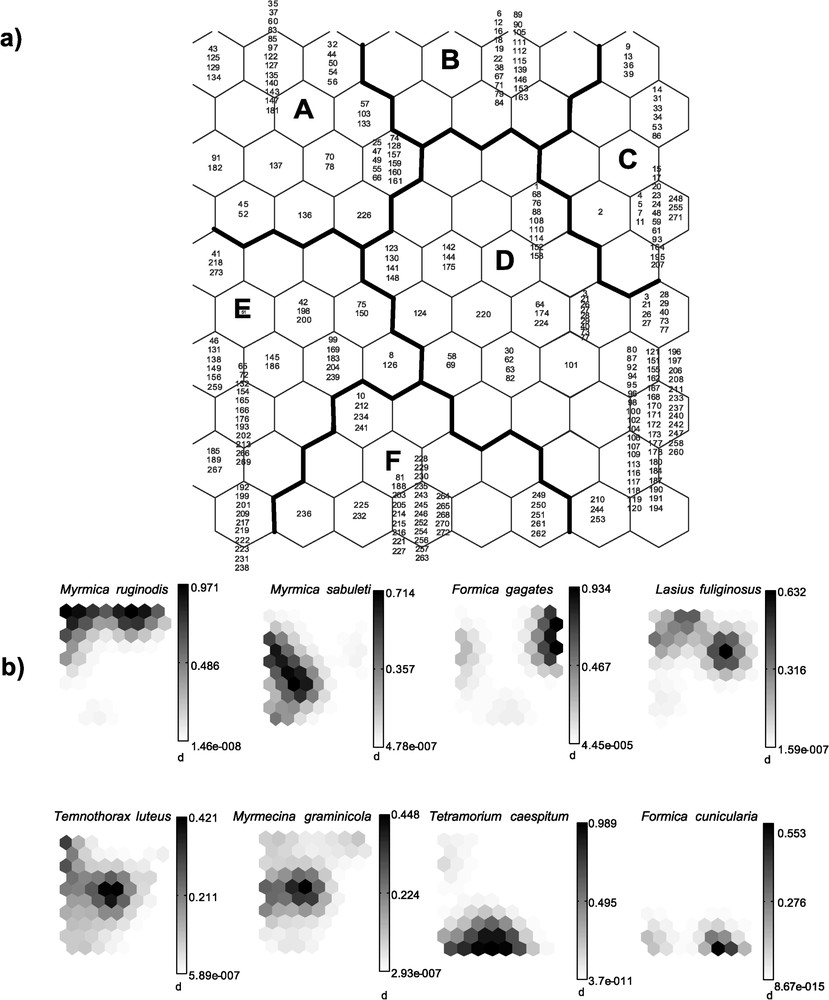
(a) Distribution of samples on the self-organizing map (SOM) according to the ant species, and clustering of the trained SOM. (b) Gradient analysis of density for each species on the trained SOM, represented by a shaded scale (dark=high values, light=low values). Each small map (corresponding to one species) is to compare to (or to superimpose onto) the map representing the distribution of samples presented in Fig. 3a, thus showing the occurrence probabilities for each species (in shades of grey) within each cluster.
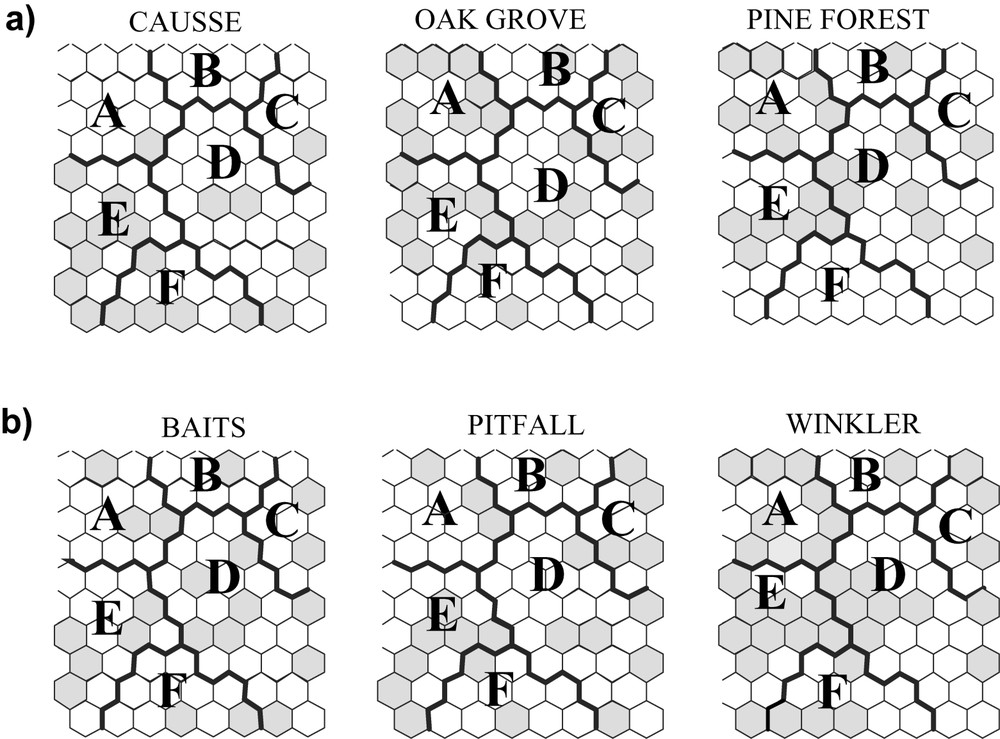
Distribution of environments (a) and sampling methods (b) on the SOM trained with ant species, represented as grey cells.
Among the most frequent species, one can distinguish those represented in only one cluster from those, more generalist, represented in two clusters or more (Appendix A; Fig. 3a and b), such as Myrmica sabuleti or M. ruginodis. The species represented in one cluster also characterize a vegetal formation (bio-indicators): Formica cunicularia and Tetramorium caespitum for the causse; F. gagates for the oak grove; and Lasius fuliginosus, Temnothorax luteus and Myrmecina graminicola for the pine forest. Other less frequent species are associated with these numerically dominant ants, thus constituting species assemblages more or less specific to each vegetal formation; their distribution is ‘influenced’ by the environment.
When the distribution of environments and sampling techniques was represented on the SOM previously trained with ant species (Fig. 4), all environments and techniques were present in the six clusters. The causse is mostly characterized by the cluster F (Tetramorium caespitum, the most frequent species), while clusters A and B (Myrmica ruginodis) characterize both the oak grove and the pine forest, and cluster C (Formica gagates) permits the oak grove to be differentiated from the pine forest. Concerning the sampling techniques, cluster A rather characterizes the Winkler extractors, the group constituted by clusters B, C and D rather characterizes pitfall traps, while baiting samples are dispersed throughout the entire chart, but are greater for cluster F. We therefore noted both an ‘environmental influence’ and a ‘sampling method influence’, so that the sampling can be biased and it is more correct to speak of ‘species assemblages’ than of ‘communities of species’ to characterize the set of ant species recorded in each vegetal formation.
Significant differences between the clusters represented in the SOM were found for the environment (; ) and the sampling technique (; ) (Fig. 5). Simplifying the models made it possible to reduce the number of factors for each variable. Five groups were obtained for the environment (; ) by grouping two clusters (A and B), and only three groups for the sampling methods (A; ; ; ; ). These simplified models are not statistically different from the corresponding complete models (; for the environment; ; for the methods).

Sample percentages according to the environment (e.g., vegetal formations) or the sampling methods for each SOM cluster. Clusters with different letters have significantly different distributions (P<0.05) for their environments (left-hand figure) or for their sampling methods (right-hand figure).
4 Discussion
The present study demonstrates that the myrmecofauna of the three compared vegetal formations is composed of several associations of more or less specific ant species. Nevertheless, certain species appear to be exclusive to a type of vegetal formation and can be key species in ant communities, and thus play a major role in the structure of these communities and likely in the rest of the environment [24]. This is the case for F. cunicularia and T. caespitum in the causse, F. gagates in the oak grove, and T. luteus and M. graminicola in the pine forest. In addition, the myrmecofauna in the pine forest is undoubtedly made up of a greater number of generalist species than in the causse or oak grove where several species are likely to occupy specialized niches. For instance, Aphaenogaster spp., Messor structor and Plagiolepis pygmaea, all absent from the pine forest, were reported to be adapted to certain soil conditions and variations in seed and prey availability [25].
Each sampling method applied to a vegetal formation is subject to biases due to practical limitations and differences in behaviour between species, and so it has its own advantages and disadvantages. The efficiency with which any species is captured by any particular technique depends on several factors, the most significant being population densities, nest distribution, foraging strategy, and body size [26]. In addition, differences in the probability of capture between species can result from certain species avoiding the trap or escaping [27]. Differences in the ants' level of activity can be affected by the period of the year, itself related to their physiological state and foraging behaviour [28,29]. Habitat structure can also influence a sampling technique's probability of capture because of either the influence of a microclimate or the habitat's physical characteristics [30].
The four techniques used in this study differ in their effectiveness and their reliability in estimating the ant species diversity of the compared vegetal formations. Even if a sampling effort is sufficient, none of these techniques will lead to the capture of all of the species because each targets a certain category of ants and thus favours their capture. We concur with a previous study stating that the use of litter extractors plus pitfall traps was recommended to study the ant fauna of temperate forests [11], and we support the idea that the Winkler extractor should especially be included in all protocols aimed at inventorying ground-dwelling ants, whether they are from tropical or temperate areas [8]. Moreover, sampling with Winkler extractors is an alternative method of collection for both epigaeic and hypogaeic ants, and the use of this technique can make the use of pitfall traps redundant for sampling epigaeic ants when litter abounds, as it does in forests [31]. Pitfall traps, which permit several sites to be sampled simultaneously [7], are more productive in open environments such as savannas and meadows (and the causse in this study) because ant capture can be compromised by the complexity of the habitat (including the density of the vegetation, the roughness of the soil surface, or the spatial arrangement of microlandscape features) [30–32]. Nevertheless, the distribution of ant nests and differences in the probability of capturing according to species can be biased with this technique [26]. These arguments added to our results highlight the need to use a combination of techniques to obtain an exhaustive sampling of the temperate ant fauna, just like in tropical areas.
Finally, several reasons might explain why the rarefaction curves did not reach their horizontal asymptote. A greater sampling effort may have permitted this to happen, but it would have been very costly. Our experience has shown that this ideal objective remains elusive, since in ant communities (except in a very simplified habitat), even a large sampling effort never allows the entire ant community to be collected, as there are always rare species that are not caught (see [5,33,34]). In addition, finding rare species is not necessarily related to a greater sampling effort, not to mention that the more an environment is sampled, the more ‘tourist species’ (sensu Belshaw and Bolton) [35] are caught. The inclusion of rare and ‘tourist’ species in such analyses explains why it is so difficult for the rarefaction curves to reach saturation.
Although studies conducted in both tropical and temperate climates are characterized by this, two main differences distinguish them. Firstly, the diversity of tropical ant species (among other things) is by far superior to that in temperate climates [32–36]. Secondly, the ant species turnover is very high in the tropics, while nearly non-existent in temperate areas [36,37]. These two factors explain why the sampling effort should be much greater in the tropics to obtain an exhaustive inventory of the ant fauna, but this is more expensive and time-consuming. Fortunately, the complementarity of the sampling methods reduces the effort necessary while providing the most complete information on the ant community.
To conclude, the protocol used to conduct inventories of tropical ground-dwelling ant fauna is also applicable to temperate areas, since the combination of sampling methods tested here seems to be well adapted to sampling temperate ants from both open and forested environments. However, with a view to obtaining an exhaustive inventory, we should increase the sampling effort and improve the protocol. In future studies on these types of environments, it would be necessary to sample several causse, oak groves, and pine forests to know better the local and regional ant fauna. It would also permit us to compare the turnover of tropical and temperate species, which would contribute to a better knowledge of the differences between the structures of tropical and temperate ant communities.
Acknowledgements
This research was made possible thanks to Dr. Raynal and the ‘Conseil général de l'Aveyron’ (logistical assistance and financial support). We are grateful to the Micropolis team for its organizational and technical assistance, Andrea Dejean for proofreading the manuscript and Marc Gibernau for his help and advice on the statistical analyses. This study was carried out while J.H.C. Delabie was invited by the ‘Université Paul-Sabatier’, Toulouse, France, as Professeur visiteur.