

1 Introduction
Choosing a new habitat suitable for living and reproducing is an important step in the settlement process of animals: it strongly influences both the spatial distribution and dispersal strategies within populations [1]. In some cases, this choice can be influenced by the presence of conspecifics [2]. Indeed, conspecific attraction (or retention) is found in various taxonomic groups throughout the animal kingdom [3].
The mechanisms used to assess the number of conspecifics in a given patch depend to a great extent on the type of information available to the animal [4]. Chemical signaling (or marking) is common across animal taxa, and is used for communication in a wide variety of behavioural contexts [5–9]. In silk-spinning arthropods, such as spiders, silken threads can also be indicative of the presence of conspecifics, and can therefore be used as a social cue for group cohesion or for selection of a new habitat [10–12].
Within a population, individuals can have the ability to identify and to respond differentially to conspecifics depending on their level of genetic relatedness, a cognitive process called kin recognition [13]. Behavioural discrimination may in some cases, follow recognition [14,15]. Genetic relatedness can therefore play a key role in the organization and functioning of social groups. The existence of kin recognition has now been demonstrated in a wide variety of taxa (e.g. mammals, fishes, insects, spiders, mites) [16–21].
Tetranychus urticae (also called the two-spotted spider mite) is a phytophagous mite that lives in extremely large groups. One of the characteristics of this mite is its abundant silk production [22]. A dense collection of silken threads (called webbing or web) is deposited over infested foliage and provides many advantages to the mites (i.e. communication, dispersal, protection against abiotic and biotic agents) [23–26]. Previous studies have suggested kin recognition in T. urticae during mate choice (i.e. inbreeding avoidance) [27] and egg-laying behaviour (i.e. modification of the offspring's sex-ratio) [19]. Although this species is composed of an assemblage of strains [28,29] no study has yet been made about the role of silk in group cohesion and strain discrimination. A preferential settlement with closely related mites would make the web sharing much more profitable (to both their direct and indirect fitness) than a web sharing between unrelated. Indeed, living with related individuals becomes advantageous when considering the time and the energy invested in the production of silk (i.e. by reducing the per capita cost of producing protein-rich silken threads) [30,31]. Perception of genetic relatedness could have evolved in the context of web sharing and avoiding intrastrain competition.
Recent studies have shown that the silk seems to be a cue for habitat selection [26,32]; the next step should be to investigate whether the mites can discriminate and alter their decision to settle when faced with silk spun either by an individual from the same strain or from another one. Since strain recognition of silk is known to alter the spatial organization of spiders [33,34], one may wonder whether changes – even slight ones – of silk between strains of mites may participate into their spatial and/or temporal distribution.
Our aim was to determine if the silk spun by one strain of T. urticae can be recognized and used by another strain as a communication cue for settlement choice. Therefore, we used two genetically different strains of spider mites (i.e. collected in different geographical localities). We compared the response of individuals from the two different strains in the presence of silken threads spun by mites from the same or from the other strain.
2 Materials and methods
2.1 Mite strain
Two strains of T. urticae were used in this study: URT (Louvain-la-Neuve, Belgium); and CAR (Tunis, Tunisia). URT was collected in October 2005 from greenhouse-grown banana leaves (Musa sp.), and CAR was collected in January 2006 from orchard-grown citrus leaves (Citrus sinensis). They had distinctive genetic backgrounds with a FST value (pair-wise differentiation test for population differentiation using allelic distribution, GENEPOP) [35,36] of 0.46 (χ2 = 132.85, DF = 10, P < 0.001) [37].
2.2 Rearing conditions
During the last 6 years, these two strains developed under the same laboratory conditions. They were reared on bean leaves (Phaseolus vulgaris L.), a preferred host plant. Leaves were placed on moistened cotton in Petri dishes (85 mm in diameter, 13 mm deep). Petri dishes were placed in boxes and kept in a climate room maintained at a temperature of 26 °C, a relative humidity of 50–60% and a photoperiod of 16 hours with 8 hours of dark (L16:D8).
2.3 Influence of related silken threads on the spatial location of mites
All individuals used in this study were one-day-old adult females. Using the same experimental design as [26], we examined whether adult females preferred to settle on a clean or a silk-covered area (spun by a female from the same strain) of a bean leaf. A 10 × 20 mm flat section of bean leaf (Fig. 1a) was divided into two equal areas (leaf side) by a piece of wet paper (1 mm in width). In order to create a silk-covered part, a female (called a weaver female) was introduced onto a randomly selected leaf side for 1 hour. During this time the weaver female did not lay (or very few) faeces or eggs. For our experiments, we exclusively used set-ups free of any external deposits except the silk. The weaver female and the wet paper dividing the two sides were then removed. Five minutes later (once the leaf surface had dried), a new female (called a test female) was placed on the rectangular bean leaf section. After 3 hours, the side of the leaf (clean or silk-covered) on which the test female had settled was recorded (n = 74 for the CAR strain, n = 63 for the URT strain). Preliminary observations confirmed that all females settled on a leaf side within the 3 hours (n = 30). Moreover, pretests were made with brushed set-ups (i.e. removing the silk spun by the weaver female) to confirm that the factor influencing the settlement of the tested mites was actually the silk (n = 30).

Experimental design for testing individual preferences: a: same strain silk; b: another strain silk; c: when same and another strain silken threads are present on a leaf surface. Colours (white and black) indicate the two strains used (white-URT and black-CAR).
2.4 Influence of unrelated silken threads on the spatial location of mites
The same design as described above was repeated, except that for each replicate, the weaver female and the test female were not from the same strain (Fig. 1b). Test females from the CAR strain were exposed to silken threads spun by URT females, and vice versa. Female choice of leaf side was recorded after 3 hours (n = 60 for CAR strain and n = 50 for URT strain).
2.5 Choice test of mites between related and unrelated silken threads
Again with the same experimental design, we introduced two weaver females (one from the CAR strain and one from the URT strain) to the bean leaf section, each to a different side. After removing the weaver females and the wet paper, a test female (CAR or URT) was placed on the leaf and its choice of a location was recorded after 3 hours (n = 50 for both the CAR and URT test females, Fig. 1c).
2.6 Data analysis
Fisher's exact test was used to determine whether settlement decisions differed between strains for each experiment (related silk/unrelated silk/choice test). When strains did react the same way to the presence of silk, data from both strains were pooled and experimental results were analysed using binomial tests (with the null hypothesis being that an individual would choose each of the two sides with a probability of 0.5). Holm's tests were used to correct the P-values obtained in each experiment. Tests were performed using GraphPad Prism version 5.01 for Windows (GraphPad Software, San Diego, California, USA; http://www.graphpad.com).
3 Results
3.1 Influence of the presence of related silken threads on the spatial location of mites
The CAR and URT strains were similarly influenced by the presence of related silken threads (P = 0.743, Fisher's exact test comparing the two strains). Individuals from CAR and URT strains had a higher probability of settling on the side of the leaf covered by silken threads (P = 0.001836, n = 137; binomial test). Fig. 2 shows the percentage of females choosing the “clean side” (white box) or the “silk side” (grey box) of the bean leaf.
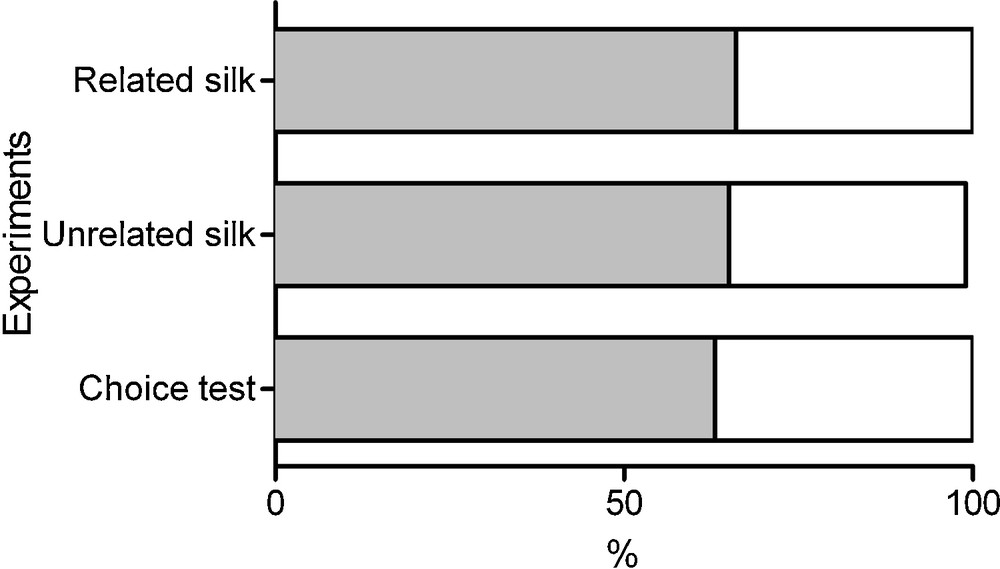
Influence of the silk on the settlement of mites (pooled data, individuals from both strains). Individuals had a higher probability of settling on the side of the leaf covered by silken threads (P = 0.001836, n = 137; binomial test). This preference did not differ according to the silk origin (same/another strain) (P = 0.944 for CAR, P = 1.000 for URT, Fisher's exact test comparing the data with related or unrelated silk side versus clean side). When individuals were confronted to both own and another strain silk, females preferred to settle in the leaf side with related silken threads (P = 0.0408, n = 100; binomial test).
3.2 Influence of the presence of unrelated silken threads on the spatial location of mites
To analyze if strains (CAR or URT) settled more often on the webbed area spun by a CAR (or URT) individual than on the one spun by a URT (or CAR) individual, we compared the data obtained in the experiment with related silk side versus clean side and data obtained with unrelated silk side versus clean side. Settlement decision for both strains did not significantly differed according to the origin of the silk (i.e. same or another strain silk; CAR, P = 0.944 and URT, P = 1.000, Fisher's exact test comparing the settlement data when silk was spun by a same or another strain).
3.3 Influence of related and unrelated silken threads on the spatial location of mites
The level of preference for silk spun by their own strain was similar between CAR and URT (P = 1.000, Fisher's exact test comparing the two strains). Females showed a higher probability of settling on the leaf side with silk spun by a female of their own strain than with the silk from the other strain (P = 0.0408, n = 100; binomial test, Fig. 2).
4 Discussion
Living in group with genetically related individuals may confer an advantage greater than its cost measured in terms of inclusive fitness [38]. In the case of two-spotted spider mites, living with individuals from the same strain could be advantageous when considering the time and the energy invested in the production of silk [30,31]. Indeed, because silk is composed mainly of proteins [30], it is expected that spider mites allocate considerable resources to produce it [39]. One potential benefit of strain recognition is the production of an expensive resource (silk) for the benefit of genetically close individuals (e.g. protection and hatching of related eggs) [40]. In addition, T. urticae individuals build a collective and protective web by weaving silken threads, cooperating in its enlargement and repair [41]. Being in a group may be favoured when the per capita costs of silk production are high making thus web sharing highly beneficial. The web seems to be a prevailing factor inducing the formation of mite aggregates, when observed in the field from leaf to leaf or plant to plant [42,43]. Aggregation may help individuals avoid predators by diluting each individual's risk of being attacked (dilution hypothesis) [44].
As silk is a key element for the protection, aggregation, group dispersal and communication of two-spotted spider mites [23,24,26,32,37], we hypothesized that the settlement behaviour of individuals (from two strains) could be influenced by the origin of the silk. Moreover, we investigated whether the silk could be involved – at least partially – in strain discrimination within T. urticae complex. Our results showed that the silk is used as a cue for settlement and strain recognition.
Perception of genetic relatedness is not uncommon in mites and it has been shown to influence behavioural decisions [45]. For example, females of the predatory mite species Iphiseius degenerans prefer to oviposit near closely related conspecific eggs rather than near eggs of more distantly related females [46]. Also, when given a choice, spider mite males preferred to guard last-moult stages of females collected from the same host plant than females collected from another host plant [47]. Finally, T. urticae females have the ability to assess mate relatedness choice as a mechanism to avoid inbreeding [27].
Strain recognition and discrimination between silk could be due to differences in the chemical signature of silken threads. Chemical signaling is common across animal taxa, and is used for communication in a wide variety of behavioural contexts (e.g. species identity, age, sex) [5–9]. As attractiveness of T. urticae silk-trails decline with time, trail-bound pheromones could be involved in settlement decisions of spider mites [26]. Differences in pheromone quantity, quality and nature between the two strains could explain our results. Further work on the chemical compounds of silk is needed to better understand how they vary between strains, stages, or sexes and how they influence communication between mite individuals belonging to same or different strains.
Results obtained in the first experiment (influence of related silk) support the idea that the two-spotted spider mite is a group living species benefiting from the presence of conspecifics (i.e. Allee effect) [48,49]. Indeed, group living brings advantages for both the individual and the group as it provides easier access to food and mates as well as protection against predators, collective modification or amelioration of the environment [50]. In T. urticae, silken threads orient not only the dispersal [23,26] but also influence the spatial location of mites. The presence of conspecifics – through the laying of silken threads – could therefore have important implications in metapopulation dynamics by shaping the settlement behaviour of T. urticae [51]. Moreover, the silk of unrelated individuals seems to influence the mites during their settlement decisions. When mites colonize a new habitat where unrelated silk is already present, it seems better to settle on a silk-covered area (whatever the silk origin). This could explain why mites (from different strains or species) could aggregate on the same host plant in more natural conditions (such as greenhouses), or in the same silk-ball formed by hundreds of dispersing mites [23].
Because T. urticae individuals quickly exhaust the host plant, they recurrently disperse to new host resources. Indeed, mites regularly move through different modes of dispersal (e.g. ambulatory, aerial, phoresy) [52]. Moreover, the trading of ornamental plants, fruits and vegetables increases the probability that strains (or even species) of mites from one country may settle or invade another location.
Strain recognition observed in T. urticae could result as a possible by-product of a finer kin recognition system existing in this species [19]. The use of silk cues would certainly make sense for a spider mite, which can only assess its own level of protection by settling down at a site. Webs could provide chemical and/or physical cues indicating where to settle (strain discrimination) and therefore where to find appropriate conditions (such as food, habitat, or mate) for living. Future experiments should investigate to which extent a group of mites (from both strains) are also able to discriminate the origin of silk so that they will preferentially aggregate with related individuals.
Disclosure of interest
The authors declare that they have no conflicts of interest concerning this article.
Acknowledgments
We thank Dr Aina Astudillo Fernandez and Dr Gilles San Martin for their help and valuable advices on the data analysis. We are grateful to Dr Lebdi Grissa Kaouthar and Marc Migon who supplied the different T. urticae strains used in our experiments. We also thank Dr Ellyn Bitume for her useful discussions on T. urticae. We are grateful to the teams from Université catholique de Louvain and Université libre de Bruxelles. This work was supported by the “Fonds pour la Formation à la Recherche dans l’Industrie et dans l’Agriculture” (FRIA) of Belgium and by the Fund for Fundamental and Collective Research (FRFC, convention 2.4622.06). This paper is number BRC216 of the Biodiversity Research Centre (Université catholique de Louvain).