

1 Introduction
In addition to direct habitat destruction, one of the main causes of human threat to biodiversity is the biological invasion of the natural and semi-natural habitats [1–4]. In recent decades, this phenomenon has been recognized as a modern research field of ecology that integrates elements of fundamental science [5] and contains numerous practical implications for the environment and environmental conservation [6,7]. For example, 1094 species on the pan-European level have documented ecological impacts, and 1347 species have economic impacts [8]. The number of publications in which authors attempt to predict the occurrence and impact of indigenous species on invasion biology is growing almost exponentially [9].
In the context of global invasive biology, vascular plants are the most researched group, containing 44% of all the invasive species studied [10]. Geographically, North, Central and South Americas and Europe have the highest proportion of species studied and are mentioned in a number of thematic published papers [10]. The first synthetic view of the alien flora of Europe [11] is obsolete and contains numerous inaccuracies in the data for individual countries [12]. Recently, a more contemporary review of the situation in the area was conducted [13,14] and the large project European Invasive Alien Species Gateway (DAISIE) resulted with more than 6000 alien plant species in Europe [13,15]. However, the lack of national inventories of alien (non-native) and invasive plants in Europe largely offsets the implementation of the DAISIE project [15]. The quantity and availability of the data are varied and often inappropriate for some parts of Europe [13,14], especially for its south-eastern (SE) area. Unfortunately, at that time Croatia was one of those poorly-recorded countries with unavailable exact data about alien and invasive flora [13]. Therefore, our intention was to contribute to the knowledge of invasive alien plants (IAS) for this part of southern Europe, where on a small geographical range, Mediterranean, Alpine and Pannonian influences “meet together”. Such variety of climate and ecological conditions resulted in a big diversity of habitats and native flora, often threatened by IAS.
Non-native and invasive Croatian flora has never been systematically treated before the year 2006 when our national survey started. Currently, there are 606 alien species and subspecies under review [16], which comprise 11.9% of the total flora, which is significantly lower than the 30% reported for some of the neighbouring countries [13]. In terms of the IAS, the standard terminology and definitions were adopted [17], and a preliminary list of invasive alien plants was developed [18]. This preliminary list is comprised of 64 taxa (Appendix A), including three subspecies of the Erigeron annuus. The greatest number of species comes from the families Asteraceae and Poaceae, followed by the family Solanaceae, while the 10 most important families are globally recognized and similarly contribute to the number of alien and invasive plants as in other countries, namely the Czech Republic, Portugal, Switzerland and Spain, but also in Europe as a whole [14,19]. Similar to the European overview [14], the life-form analysis of the invasive plants in Croatia showed the predominance of therophytes (56.3%), and the analysis of geographic origin showed that most of the invasive plants originate from the Americas (71.9%) [18].
Furthermore, a new module of the Flora Croatica Database (FCD)–Allochtonous (alien, non-native) plants was designed exclusively for monitoring, data collecting and data dissemination [16]. Thus, information for invasive alien plants in different regions of Croatia is gradually accumulating in the literature [20–33] and in six national herbarium collections (ADRZ, CNHM, MAKAR, NHMR, ZA, ZAHO) registered in the Index Herbariorum [34]. So far, there are only a few publications concerning the distributional properties and habitat preferences using small-scale surveys [35–37]. However, the accumulation of data in the FCD, digitalization of the collections and publication of the data and field observations provides the platform for the first overall insight into the spatial distribution of invasive plants in Croatia and their relation to the relief, climate and habitats. Therefore, the aims of our work were:
- • to analyse the quality of data about invasive alien plants in Croatia accumulated in the Flora Croatica Database (FCD);
- • to provide first comprehensive quantitative information on invasive alien plant taxa in Croatia useful in possible future monitoring and research projects;
- • to define the spatial level of plant invasions on this area;
- • to explore the distributional patterns of invasive alien plants regarding human population density, longitude/latitude ratio, altitude, meteorological factors and habitat types.
The results should contribute to the better knowledge of one of the main threats to biodiversity, alien and invasive flora in both Croatia and Europe, and fill the lack of national data from southern and south-eastern Europe [13,38,39]. Furthermore, the results should provide a reliable regional and global analysis for strategic planning regarding invasive plants management and biodiversity conservation.
2 Materials and methods
2.1 Study area
Croatia is a country in south-eastern Europe (Fig. 1), comprising a land area of 56,594 km2 and a sea area of 31,067 km2. Altitudes of up to 200 m cover 53.4% of the land area, 25.6% of the area is located 200–500 m above sea level and 21% of the total land area is > 500 m above sea level. The highest peak reaches 1831 m a.s.l. (Dinara Mountain). The total length of the watercourses within the land boundaries are 3500 km, and the longest rivers are the Sava (518 km), Drava (323 km), Kupa (293 km), Bosut (151 km) and Danube (135 km). A total of 22,333 km2 of land area is covered by different types of forests (49.4%).

Location map of Croatia and of the places discussed in the text.
The annual mean temperature in Croatia (40-year period) ranges from 3 °C in the highest mountain area, to 17 °C along the coast and on the islands. The mean annual precipitation ranges from 300 mm to slightly over 3500 mm. The smallest annual amount of precipitation falls on the outer islands of the southern Adriatic, and the largest annual precipitation falls in the Gorski Kotar mountain region [40].
The main geological feature of Croatia is the presence of large karst areas. The Dinaric karst area (Dinaric Alps) consists predominantly of carbonate rocks of the Mesozoic age. It stretches from the Julian Alps from the Northwest and continues to the southeast Balkan Dinarids until reaching the Helenids in Greece [41].
In terms of the mesoscale landscape, three relatively clearly demarcated entities can be identified (Fig. 1): Lowland Croatia bordered by the Sava, Mura, Drava and Danube rivers is dominated by the wide areas of oak-woods, marshes, wet meadows and pastures; Highland Croatia contains karst mountain strip forms to pre-stretch mountains, and a section of the Dinara mountain region is dominated by forests of fir and beaches, with diverse geomorphology (> 8000 caves, pits, ravines and karst valleys); Mediterranean Croatia belongs to the eastern Adriatic shoreline, including the coast and the islands.
The Adriatic Sea is a deep natural bay located in the central part of the Mediterranean that presents the most northern protuberance of the Mediterranean in the European continent, which is situated between the Apennine and Balkan peninsulas and surrounded by the mountain ranges of the Apennines, the Alps, Dinarids and Helenids. The Adriatic basin follows the Northwest-Southeast (NW-SE) direction similar to the Dinaric Alps, for a total length of 783 km. The eastern Croatian coast is one of the most diverse in the Mediterranean area [42], with high biodiversity [43].
2.2 Data analysis
The selection of invasive vascular plant taxa was based on the upgraded online inventory of Croatian vascular flora in the Flora Croatica Database [44], and follows a preliminary list of 64 invasive alien plants [18] (Appendix A). For these taxa, all of the available spatial information was accumulated in the Flora Croatica Database [44]. The geocoded findings originated from three sources: the literature, herbarium collections (ZA, ZAHO, CNHM) (abbreviations according to [34]), and field observations performed from 2004 to 2012.
The spatial analysis was conducted using several different spatial units:
- • MTB¼ network for mapping the flora of Central Europe [45], which was extended to Croatian territory as the national standard [46]. Each grid cell of this network has an area of approximately 35 km2 (5′ × 3′). There are a total of 2028 grid cells. This grid was used to display the spatial and data distribution of the number of species per grid cell and to estimate the correlation between the number of invasive plants with the longitude/latitude ratio;
- • areas of the 6749 settlements [47] for the relation of the human population density with the number of recorded invasive species within;
- • basic grid cell of 1 km2 for analyses of invasive plants dependence on altitude and on environmental variables; namely: mean annual temperature; precipitation, insolation duration and global solar irradiation. The same grid was used to determine the preference of invasive plants to particular habitat types at the first and second level of the Corine land cover (CLC) classification [48].
The data on the relief were obtained from the Digital Elevation Model (DEM) of Croatia developed from the topographic maps in scale (1:25,000) with the available basic information for 100 × 100 m DEM cells. The adjoining mean values of all DEM cells that fall in each 1 km2 grid cell were used as corresponding values for elevation. The data on climate were derived from [40] based on measurements from 700 meteorological stations for the period of 1961–2000, and climate models were constructed for the whole area [49–51]. The climatic variables used were mean annual temperature (°C), mean annual precipitation (mm), insolation duration (h) and mean annual global solar irradiation (Jm−2). Due to equal spatial resolution, i.e., 1 km2, the values were assigned on a cell-to-cell basis.
The data concerning habitats were obtained from the Corine land cover maps from 2000 [52] and from the Map of Habitats of the Republic of Croatia (1:100,000 scale)–State institute for Nature Protection [48]. Habitat diversity is expressed as a Shannon–Wiener index of diversity (H) [53]:
The calculation per selected spatial unit was performed using BioDiv ver. 1.1 (developed by Faculty of Science and Faculty of Electrical Engineering and Computing) and the Flora Croatica Database [44]. The GIS evaluation was conducted using ArcView 3.2a and ArcMap 10 [54]. The statistical analysis was conducted using Statistica 8.0. Software [55].
3 Results
According to our results, Croatian flora has been researched since the 18th century, but remarkable progress in invasive plants findings has started from the middle of the 20th century (Fig. 2A), which resulted with an increase of new findings of invasive taxa. There are almost 6000 data from field observations, a little bit less literature data, and a smaller amount of data from herbaria (ZA, ZAHO, CHMZ) related to the problem of IAS and accumulated into the FCD (Fig. 2B). In recent decades, the accuracy of the data is also increasing in terms of geo-precision (r = 0.42, P < 0.05; Fig. 2C). The first mention of invasive species in the FCD comes from literature published at the beginning of the 19th century [56] concerning Sorghum halepense (noticed for the islands of Dalmatia) and Xanthium spinosum (noticed for the city of Koper, Slovenia). The data about spatial distribution of invasive plants are not homogenous, because the flora of Croatia is still not completely mapped. The best researched are protected areas, but there are still some smaller areas without floristic data.
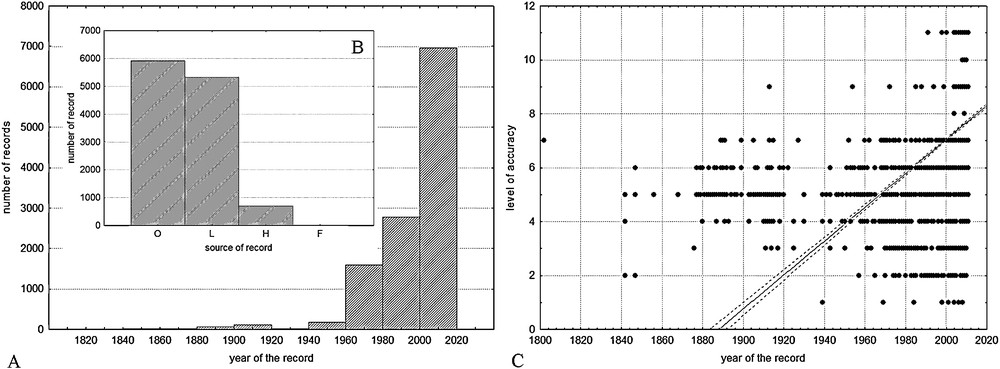
The accumulation and accuracy of IAS distribution data. A. The dynamics of the accumulation of data regarding invasive species. B. Data shared from different sources (O: field observation, L: published data, H: data from herbarium collections, F: geocoded photo documentation). C. Dependence of the level of accuracy on the period of data collection (y = −118.6877 + 0.0629 × x; r = 0.42, P < 0.05).
Our study of invasive alien plants in Croatia included 64 taxa (Appendix A). We found out that they occur in a total of 989 MTB¼, which comprises 48.76% of all MTB¼ Croatian fields and covers 34,615 km2 of the state territory. On average, most of the grid cells (each grid cell is approximately 35 km2) contained 4.7 invasive species, with a minimum of one, and in particular cases, up to 47. Their spatial distribution is expectedly not uniform (Fig. 3). The greatest number of invasive species (> 30 per grid cell) was recorded in the major urban centers (primarily in Zagreb and Split areas) (Figs. 1 and 3). Up to 20 species per grid cell are characteristic for smaller urban centers, such as Osijek, Rijeka, Makarska and Dubrovnik (Figs. 1 and 3). Up to 10 invasive species per grid cell are primarily characteristic for the major river valleys, such as the Sava, the Drava, the Danube, the Krka and the Cetina (Figs. 1 and 3). Areas with a high diversity of invasive plants are actually relatively small and limited to the mentioned urban areas (Figs. 1 and 3). Anthropogenic influence on the occurrence and spreading of invasive plants is also confirmed by the positive correlation between the population in the 6749 settlements and the number of invasive species in those urban areas (r = 0.367, P < 0.05; Fig. 4). The largest area in which invasive species occur is characterized by the presence of a small number of those species (Fig. 5A). The largest number of grid cells (712) actually contained less than six invasive species; thus, invasive species diversity is relatively low (Fig. 5A).
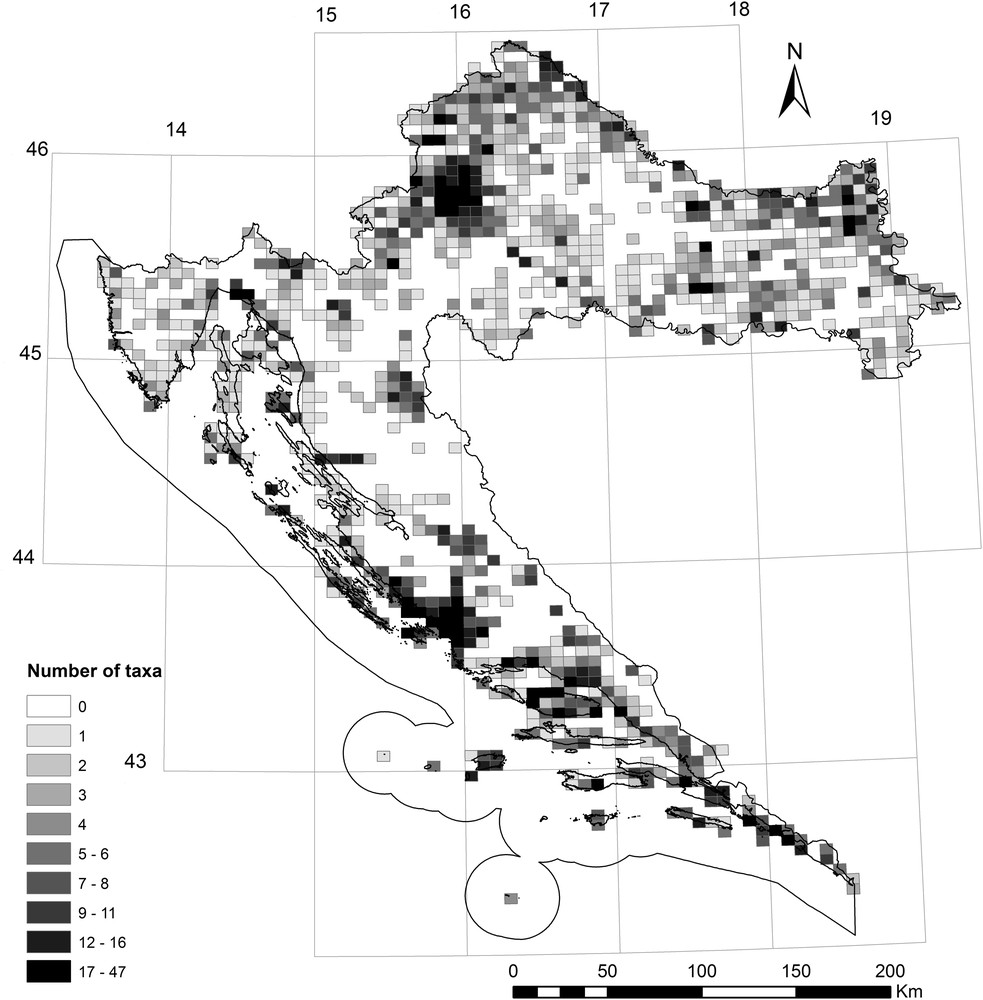
Map showing the distribution and number of invasive species and subspecies per grid cell (MTB¼; approximately 35 km2) in Croatia (classified into 10 classes, with the delineation of the classes as 10% percentiles).
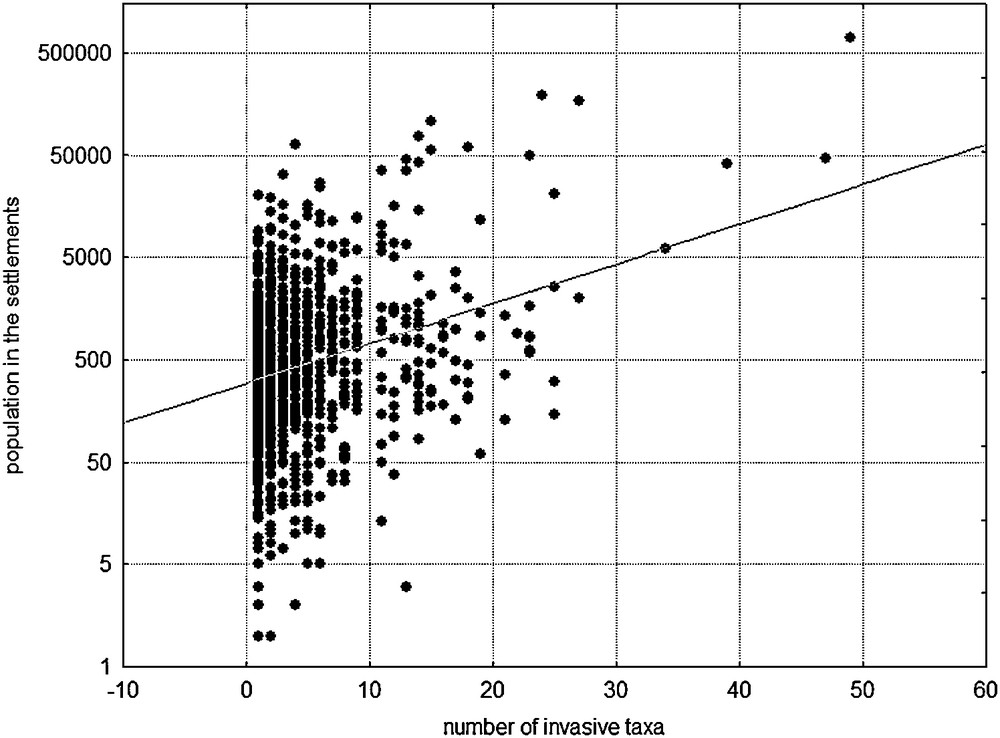
Relation between the population in the 6749 settlements and the number of recorded invasive species within the settlement areas (y = −3765 + 1609.2 × x, r = 0.367, P < 0.05).

Overall IAS distribution and longitude-latitude relation. A. Number of occupied grid cells (MTB¼) with invasive taxa in Croatia. B. Relationship between the number of invasive plants and the longitude–latitude ratio (Gauss-Krüger coordinate system): higher values are observed in SE Croatia, and lower values are observed in NW Croatia (based on MTB¼ grid cells); the number of invasive taxa per grid cell increased in the SE direction (x/y = 1.1234 + 0.0070 × number of taxa; 0.95 Conf. Int., r = 0.126, significant on P < 0.05).
The correlation between the invasive species number and the geographical coordinates ratio (longitude/latitude ratio) for each centroid grid cell indicates that the number of invasive plants in Croatia gradually grows in the southeast (SE) direction (r = 1.26, P < 0.05) (Fig. 5B).
The distribution of invasive species due to the altitude showed markedly left asymmetry (Fig. 6A). The average altitude of all of the findings was 248.5 m a.s.l. (Sx ± 4.69); and the invasive plants were the most common at altitudes from 0 up to 300–400 m a.s.l. Areas with a 400–1000 m a.s.l. had a smaller number of invasive plants and at higher altitudes (> 1100 m) the number of invasive plants rapidly decreased with increasing altitude. The correlation between the number of invasive species and the altitude is therefore negative (r = −0.137, P = 0.05) (Fig. 6B).
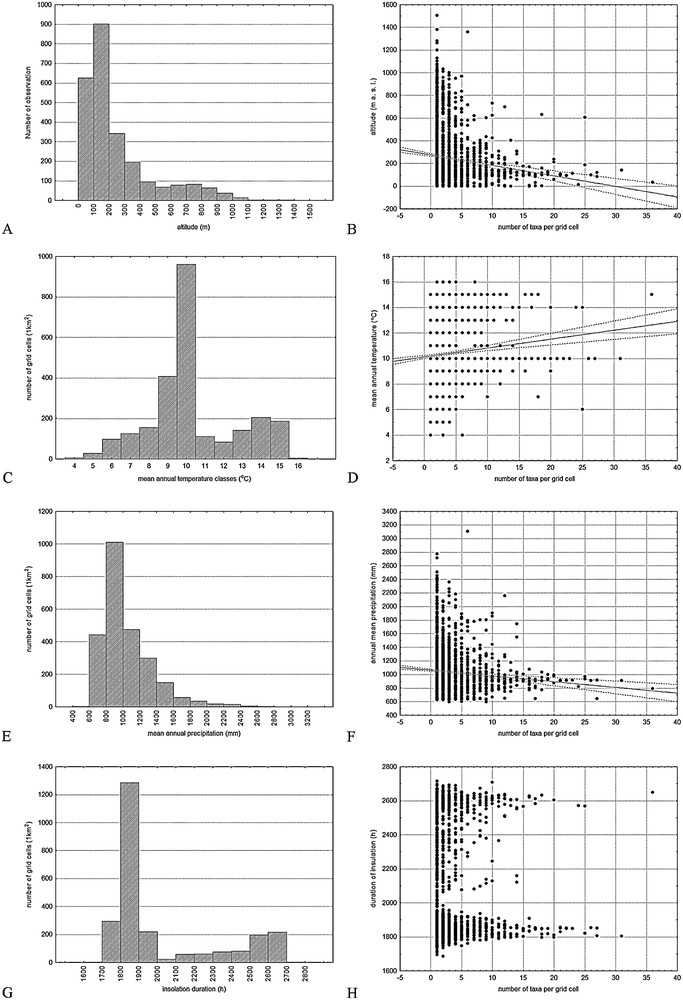
IAS distribution and relations to the abiotic factors. A. Altitudinal distribution of invasive plants occurring in 100 m altitudinal belts. B. Correlation between the number of invasive taxa and the altitude (y = 275.60 − 9.2393 × x; r = −0.137, P < 0.05). C. Distribution of invasive plant taxa in mean annual temperature classes (°C). D. Correlation between the number of taxa per grid cell and the mean annual temperature (y = 10.1247 + 0.0698 × x; r = 0.102; P < 0.05). E. Distribution of invasive plant taxa in annual mean precipitation classes (mm). F. Correlation between the number of taxa per grid cell and the mean annual precipitation (mm) (y = 1062.60 − 8.4211 × x; r = −0.095; P < 0.05). G. Distribution of the invasive plant taxa in insulation duration classes. H. Correlation between the number of taxa per grid cell and the insulation duration.
Meteorological parameters showed several regularities connected with the distribution of invasive alien plants (Fig. 6C–H). The average mean annual air temperature for all Croatian areas with invasive species occurrence was 10.33 °C (Sx = 0.04; minimum 4°, maximum 16 °C) and the distribution of invasive plant taxa in mean annual temperature classes was bimodal (Fig. 6C). The largest part of the area in which IAS appeared had a mean annual air temperature between 9 °C and 10 °C, and a smaller part was characterized by annual temperature averages between 14 °C and 15 °C (Fig. 6C). The correlation between the number of species and the average annual temperature was positive (r = 0.102; P < 0.05) (Fig. 6D). The average annual precipitation for all areas with invasive plant occurrence was 1038 mm (Sx = 6.17; min = 606 mm, max = 3107 mm) (Fig. 6E). Considering the mean annual precipitation, the distribution of invasive plants showed extreme left asymmetry (Fig. 6E) with a higher incidence of species in areas with precipitation in the range of 600–1400 mm. The occurrence of invasive plants was rare in the areas with more than 1800–2000 mm of precipitation per year. The correlation between the number of species and the average annual precipitation was negative (r = −0.095; P < 0.05) (Fig. 6F). The average insolation duration for the areas with invasive plants was 2025 h (Sx = 5.97; min = 1686 h, max = 2715 h). The distribution was bimodal, highly asymmetric to the left, with higher values between 1700 and 2000 h/year and between 2400 and 2700 h/year (Fig. 6G). The correlation between the number of species per grid cell and the insolation duration clearly showed optimums of insolation duration for the invasive species of 1900 and 2600 h/year (Fig. 6H). A similar distribution and correlation was observed for solar irradiation (Jm−2) and both of these parameters were logical, expected and related to the mean bimodal annual air temperature to which the duration of insolation and global solar irradiation were significantly correlated (r = 0.86 and r = 0.83 at P < 0.05).
With respect to invasive plant species preferences for particular habitat types, we present a graphical overview of the analyses per type from the second level of Corine land cover (CLC) classification and compare them in the following text at both the first and second levels of CLC classification [48] (Fig. 7A). The most diverse invasive flora was developed within the agricultural area (Fig. 7A) with a total of 60 invasive taxa. The average number of invasive plants in a single agricultural field was 2.8 taxa. The diversity of invasive flora was also very high within the artificial surfaces (59 taxa) (Fig. 7A). The average number of invasive plants in a particular artificial area was 4.6 taxa. A total of 56 invasive taxa were recorded in the forest and semi-natural areas (Fig. 7A), with 2.5 species on average per single land cover type polygon. A total of 37 taxa were recorded in inland waters or in the close vicinity of inland waters, and 20 taxa occurred inside inland wetlands with a high average number of species per single inland water locality (5.1) (Fig. 7A).

IAS distribution and habitat types. A. Distribution of invasive plant taxa in areas with different land cover types (according to the Corine land cover classification level 2: 11, Urban fabric; 12, Industrial, commercial and transport units; 13, Mine, dump and construction sites; 14, Artificial, non-agricultural vegetated areas; 21, Arable land; 22, Permanent crops; 23, Pastures; 24, Heterogeneous agricultural areas; 31, Forests; 32, Scrub and/or herbaceous vegetation associations; 33, Open spaces with little or no vegetation) based on 1-km2 grid cells. B. Relationship between the number of invasive plant taxa and the habitat diversity presented as Shannon–Wiener indices (y = 1.3514 + 0.01286 × x; r = 0.2, P < 0.05).
The number of invasive taxa in Croatia per grid cell increased with habitat diversity, i.e., the number of these taxa is positively correlated (r = 0.2, P < 0.05) with the Shannon-Wiener index of habitat diversity (Fig. 7B). Additionally, the correlation between the number of invasive plant taxa and the total number of habitat polygons (as a simple measure of habitat fragmentation) was positive (r = 0.278, P < 0.05).
4 Discussion
Despite the fact that the Croatian invasive flora have been more or less researched since the 18th century and there are lots of data about non-native and invasive plants in Croatia (Fig. 2) [16,44], those data are not homogeneously distributed, particularly considering the year of the recording, the accuracy of precise geographical data and spatial coverage. These factors could affect our results to a certain extent. A significant increase in IAS findings began in the early 1960s (Fig. 2A). Unpublished findings in the field dominate in the FCD, followed by literature data and a smaller amount of data from herbaria collections (Fig. 2B). Although in recent decades the accuracy of the data is increasing in terms of geo-precision, the amount of inaccurate statements in recently published literature and collected herbarium samples is unfortunately still surprisingly high (Fig. 2C). Nevertheless, the importance of the information on the first appearance of species from the invasive plants list in a particular country [57,58], the quality of such information for Croatia varies considerably. In some cases, the exact date of the species introduction in Croatia, how and where it was transferred, when it was first spotted outside of the place of introduction, etc., was known [33]. In many cases, unfortunately, there was no such data, which greatly hindered or prevented the understanding of the process of invasion [59]. The first mention of invasive species in Croatia was those of S. halepense [56]. However, this plant was registered on two islands in Dalmatia (Šolta and Ugljan) with a possibility that it was present there for a longer time [60]. So, the problem with older data, either from literature or herbaria collections, is that we cannot be sure that the first information about IAS in Croatia is actually the accurate information about the first appearance of an invasive plant. Unlike some excellent projects concerning flora mapping in Europe, only parts of Croatia are completely mapped, providing a comprehensive picture only at the local level (mostly protected areas). For the rest of the area, the field data were collected unsystematically, depending on the interests of individuals, institutions, project objectives, transportation facilities, availability, etc. The influence of the level of exploration of invasive species distribution is particularly evident in the national parks of the Plitvice Lakes and Risnjak (Fig. 1). Namely, the existence of a relatively large number of invasive plants findings in these areas does not mean that these areas are the centres of diversity of invasive plants, but that their flora is thoroughly mapped when compared with their immediate neighbourhoods (Fig. 3).
During our study we found out that the number of alien species in the flora of Croatia is significantly higher than the 157 species listed by Lambdon et al. [13] or the 161 species identified in the DAISIE [15]. The list of those species for Croatia contains 606 taxa, of which 265 are naturalized [16], although this list is not exhaustive. Specifically, the number of cryptogenic taxa in Croatian flora is high and undefined, and the degrees of naturalization and causality are not sufficiently known. According to findings from other countries of Central and Western Europe, the number of introduced species is greater than 30% [14,61], and consequently, in Croatia, at least 1500 alien plants can be expected, with a naturalized share of 1/3 according to other European states [13]. With this rough current estimation, new non-native taxa are described relatively frequently, e.g. Milović et al. [33] cited 16 new neophytes recorded in only 4 years. Therefore, the current list of invasive species is under continuous revision. From the first listing [18] until now, several potential candidates for the invasive alien plants list of Croatia have been detected (i.e., Amaranthus albus L., A. blitoides S. Watson, A. deflexus L., and A. hybridus L.), but the current data are not yet sufficient for a proper assessment. The additional taxa of globally recognized invasive species were also distributed in Croatia, i.e., Mesembryanthemum crystallinum L., Heracleum mantegazzianum Sommier et Levier, Opuntia ficus-indica (L.) Miller, O. vulgaris Miller, Lupinus polyphyllus Lindl., and Physalis peruviana L. [62], but these taxa still do not show signs of invasiveness according to criteria proposed by Richardson et al. [63], which is a circumstance that will probably change in the near future. Therefore, the basis for our study was the list of 64 IAS taxa of Croatia given by [18].
According to our results, invasive alien plants in Croatia occur in almost 50% (34,615 km2) of the state territory, with an average of five invasive plants per 35 km2 (minimum of one, maximum of 47; Fig. 3). Areas with a high diversity of invasive plants are actually relatively small and limited to urban areas, acting as the centres of diversity of invasive plants. The highest concentration of IAS in the major urban centres is located at the intersection of main continental transport corridors (Zagreb, Varaždin) and seaports (Split, Šibenik) (Figs. 1 and 3). In these areas, up to 47 invasive species per grid cell have appeared. In addition, a variety of invasive species per grid cell was found in smaller urban centres (up to 20 species per grid cell; e.g. areas of the cities of Osijek, Rijeka, Dubrovnik and Makarska) and the major river valleys (up to 10 species per grid cell; e.g. banks of the rivers Sava, Drava, Danube, Krka and Cetina) (Figs. 1 and 3). In all mentioned areas with intensive human influence invasive plants could be found almost everywhere, thanks to enormous quantity of cultivated areas, industrial, ruderal and other disturbed habitats, known as easily colonized by alien plants [13,37,64]. Further clear evidence of the strong anthropogenic influence on the number of invasive plants in Croatia is also a positive correlation between this number and the population density in settlements (Fig. 4), which confirms the opinion that contributions of alien plants increased with city size and are closely associated with human activity [65,66]. However, despite the uneven spatial exploration (Fig. 2), it is obvious that some areas are more affected and some are more resistant to invasion. The largest area of Croatia is characterized by the presence of a smaller number of taxa with less than six invasive plants per grid cell. Therefore, the diversity of invasive plants seems relatively low (Figs. 3 and 5A), but this fact could be misleading. Namely, several noxious invasive plants (Appendix A) are distributed all over the Croatian area, e.g. Ambrosia artemisiifolia, which is an enormous threat to biodiversity and public health in Croatia [67] and elsewhere [68,69]. Further careful monitoring of its spreading and planning of management actions in Croatia is important for the whole Europe, which is infected with plants of Ambrosia. Unfortunately, it occurs in most of the lowland Croatia and across the Mediterranean area, as well, although in pretty smaller quantity [16,67]. So, on this example it is clear that, in spite of a small number of invasive plants in some areas of Croatia, they must be carefully and continuously monitored and if possible, eradicated or at least controlled.
Croatian state territory is laid in the NW-SE direction, roughly following the direction of the Dinaric Alps (Fig. 1), and a moderate increase in the number of invasive plant species in the SE direction, i.e., the descent toward the NW (Fig. 5B), is consistent with some of the established global trends. Such a gradient of species diversity is one of the most robust distribution trends in biogeography [70], and it was also established for naturalized species in continental Europe [71], aliens in urban flora of Central Europe [65], etc. We showed that this trend was also observed and confirmed with our results for the Croatian territory (Fig. 5B). However, the increased presence of invasive plants in Mediterranean Croatia with relation to the continental part is somewhat inconsistent with the general assumption that the European Mediterranean biogeographical region is less invaded as a relatively resistant ecosystem against recently introduced species [4,38]. In fact, the largest proportion of recorded introductions and naturalization in the last 20 years has occurred in the Mediterranean region of Croatia [31,33,72–75]. One possible cause, though difficult to prove, is a much larger movement of people, goods and services associated with rapid tourism development in this area, which generally could support undeliberate and deliberate introductions [3].
Furthermore, our research showed that a total of 43 invasive species were recorded in all explored islands of the SE Adriatic [16] (Fig. 5B), which comprises 67% of all invasive species in Croatia, although the total area of these islands barely reaches 3000 km2, i.e., only 5.3% of national territory. The islands (Fig. 1) with the largest number of identified IAS are Brač (32 taxa), Žirje (15 taxa), Vis (14 taxa), Mljet (14 taxa), Hvar (12 taxa), Rab (12 taxa) and others (with ≤ 10 species). The most widespread invasive species on the Adriatic islands, i.e., those with the largest numbers of localities, are Conyza canadensis, Amaranthus retroflexus, S. halepense, A. albus, Conyza bonariensis, Amaranthus deflexus, Bidens subalternans, and Ailanthus altissima. This result confirmed the opinion that islands tend to harbour more alien species than the equivalent mainland sites [76,77]. Causes for the increased exposure of the islands to invasive plants are not quite clear and the phenomenon that the new incoming species invade and displace native species that are well adapted to local environments is paradoxical [77]. The possible causes are still the subjects of speculation: the island's native species are naturally less resistant to disturbances in the environment; invasive plants from inland are superior with regard to evolutionary history, accumulating more appropriate genetic modifications; low native species richness on islands, etc. [70,77,78].
The distribution of invasive species in Croatia due to the altitude showed that these plants occurred in a relatively wide altitude range, from sea level to the mountain areas, but mostly no higher than 1100 m a.s.l., i.e., with increasing altitude their number quickly decreased (Fig. 6A, B). The invasive plants in Croatia were the most common at lower altitudes (up to 300–400 m a.s.l.), which corresponded with the altitude of the lowlands and the elevations of the coastal areas in Mediterranean Croatia. Areas with 400–1000 m a.s.l. with a smaller number of invasive plants corresponded with the highland Croatia. The occurrence of species located at higher altitudes (> 1300 m) was scarce and related only to several records of E. annuus, C. canadensis and Solidago gigantea on a few localities on the Velebit Mountain and the Gorski Kotar area (Fig. 1). At higher altitudes (> 1500 m), there were actually no invasive species. The altitudes with the largest concentration of invasive taxa are characteristic for the lowland and Mediterranean Croatia, where recognized urban centres with high invasive plant diversity are placed (Figs. 1 and 3). Generally, elevation richness patterns peaking from low to middle elevation ranges were already observed [79]. The numbers of species per site declined exponentially with increasing altitude in the Swiss Alps [80], Australian Alps [81], Canary Islands [66], urban centres of Central Europe [65] and it was shown for the Medvednica Mt in continental Croatia [37]. Thus, the negative correlation between the IAS records and the altitude observed in our research (Fig. 6B) is a known phenomenon, suggesting that the gradual spread of alien species after their introduction (residence time) was a major process involved in shaping the currently observed patterns [82,83]. This fact could be caused by several reasons: a progressive loss of species, and therefore, the species found at high altitudes are those with the widest ranges that also occur at low elevations; higher elevations are less populated, which means less human disturbance on habitats; meteorological factors, etc.
Meteorological parameters for all areas with invasive plant occurrence (the average mean annual air temperature, the average annual precipitation, the average insolation duration, the average solar irradiation) showed several regularities connected with the distribution of IAS plants in Croatia (Fig. 6C–H). The largest part of the area in which invasive species appeared had a mean annual air temperature between 9 and 10 °C (Fig. 6C, D), which corresponded with nearly the entire territory, including highland and lowland Croatia (Fig. 1), except the cooler high mountain and warmer coastal areas. A smaller part of the area was characterized by higher annual temperature averages, and the invasive plants occurred at a mean annual air temperature between 14 and 15 °C (Fig. 6C, D). This area coincides with the Mediterranean and the lower parts of the coast with Mediterranean influence found deep in the continental area along the river valleys of the Adriatic basin (Zrmanja, Krka, Cetina, and Neretva) (Fig. 1). Similar trend of gently growth of the number of species with increasing temperature was observed elsewhere [65]. The same areas of Croatia correspond to the distribution of invasive plants showed according to the number of taxa regarding precipitation, insolation and irradiation. Namely, the number of invasive plants per grid cell increased with increasing aridity (Fig. 6E, F). The incidence of invasive plants is the highest in areas with precipitation in the range of 600–1400 mm per year (Fig. 6E, F) and the insolation duration from 1900 and 2600 h/year (Fig. 6H). A similar distribution was observed for solar irradiation (data not presented here) and both of these parameters were logical, expected and related to the mean bimodal annual air temperature (Fig. 6C, D) to which the duration of insolation and global solar irradiation were significantly correlated (r = 0.86 and r = 0.83 at P < 0.05).
From mentioned results it is obvious that a smaller number or the complete absence of invasive species from the higher altitudes in Croatia strongly influences their climatic profiles and vice versa because the environmental optimums do not allow growth at higher altitudes. The climatic characteristics of the mountainous belt are low annual temperatures (< 7 °C), high annual precipitation (> 1600 mm), insolation between 2000 and 2300 h/year and solar irradiation 4700–5000 Jm−2 [40]. At lower altitudes, the frost-free zone from the Mediterranean side of the Dinaric Alps in Croatia (Fig. 1) is dominated by a thermophilous invasive species native to the subtropical and warm areas of Africa, Asia and America. The presence of these taxa at higher altitudes is probably limited through low temperatures (Fig. 6C, D), which has already been observed in other areas [66,84]. Among these species are the well-known invasive species of the Mediterranean: C. bonariensis, A. albus, A. retroflexus, Carpobrotus edulis, B. subalternans, etc. On the other, continental side, there are taxa that are not adapted to the higher temperatures and arid conditions of the Mediterranean area, and accordingly, there are no reported findings of those plants from the Mediterranean part of Croatia (e.g., Asclepias syriaca, Bidens frondosa, Duchesnea indica, etc.). Moreover, there is a group of taxa with broad meteorological (ecological) amplitudes that inhabit both sides of the Dinaric Alps, i.e. both continental and Mediterranean areas of Croatia. In this group, there are species that reach the highest altitudes on the Dinaric ridge, e.g., Acer negundo, A. artemisiifolia, C. canadensis, Impatiens glandulifera, etc. However, the impact of climate change on the spread of invasive species is much weaker when compared with the impact of changes in land use [39].
Further very important influence on invasive plant species is their preference for particular habitat types, which was also confirmed by our study – the main habitats dominated by invasive plants in Croatia are those developed under the higher influence of human activities (Fig. 7A). The most diverse invasive flora in Croatia was developed within the agricultural area (60 invasive taxa occurring in Croatia i.e., 91%), with almost all IAS from the List of invasive species [18]. Within this category, variability in the number of invasive alien plants between particular habitat subtypes was not so high. Invasive taxa were detected in arable lands (mark 21 on the Fig. 7A), permanent crops (mark 22 on the Fig. 7A), pastures (mark 23 on the Fig. 7A) and heterogeneous agricultural areas (mark 24 on the Fig. 7A). The diversity of invasive flora was also very high within the artificial surfaces (59 taxa, i.e., 89.4%), which were characterized by the highest variability in the number of species between subtypes. The invasive species occurred mostly on all artificial land use subtypes, but a number of taxa per single locality occurred in the mines, dumps and construction sites (mark 13 on the Fig. 7A), and the artificial, non-agricultural vegetated areas (mark 14 on the Fig. 7A, including green urban areas and sports and leisure facilities) were higher as compared with the fabric areas (mark 11 on the Fig. 7A) and industrial, commercial and transport units (mark 12 on the Fig. 7A). An average number of invasive plants was greater in particular artificial subtypes (4.6 taxa) than in a single agricultural field (2.8 taxa). Further habitat analysis showed that a lot of invasive taxa were recorded in the forest and semi-natural areas (56 taxa, i.e., 84.85%; Fig. 7A), but with a smaller average number of species per single land cover type polygon (2.5 taxa). The average number of species per single area of particular vegetation types was higher in the open spaces with little or no vegetation (mark 33 on the Fig. 7A) as compared with scrub and/or herbaceous vegetation associations (mark 32 on the Fig. 7A) or natural forest types (mark 31 on the Fig. 7A). The 37 taxa were recorded in inland waters or in the close vicinity of inland waters (i.e., 56% of all invasive taxa that occur in Croatia), and 20 taxa occurred inside inland wetlands (i.e., 30.3% of all invasive taxa that occur in Croatia) with a high average number of species per single inland water locality (5.1) (Fig. 7A).
In the European context, it has been shown that the man-made habitats (industrial areas, agricultural areas) and other strongly affected habitats (forest areas, semi-natural habitats) have the largest share of alien and invasive plants. Of all of the naturalized alien species of Europe, 64% occur in the industrial areas and 58% in the arable fields, parks and gardens. Grasslands and forest areas are also powerfully influenced, with 37% and 32% of naturalized species, respectively [14]. Our analysis of habitat types and the number of invasive species in Croatia showed a similar pattern (Fig. 7): almost 75% of all sites with invasive plants are located within a few direct anthropogenic influenced areas, such as artificial surfaces, agricultural areas and affected forests (Fig. 7).
Our results clearly show that the number of invasive taxa per grid cell increases with habitat diversity and the level of habitat fragmentation (Fig. 7B). Higher habitat diversity and fragmentation of some spatial units result in a higher overall diversity [37,64,78,85]. Thus, also in our sample (Fig. 7B), the species richness was positively correlated with the invasive taxa richness, which is a kind of paradox. This phenomenon is shown by many large-scale surveys [5,37,38,78,86,87], although in theory, more diverse communities use resources more completely and reduce their availability to potential invaders [5]. The data up to now indicate that areas with higher diversity are more open to the naturalization of alien flora [88–90]. The numbers of alien plants which will become invasive, of course, depend on the total number of species that are introduced, the number of species that have been successfully naturalized and the proportion of naturalized taxa that develop characteristics of invasiveness [4]. This issue deserves special attention everywhere, as well as in Croatia, because invasive alien plants are mostly therophytes from the families Asteraceae and Poaceae [18], which are the best adapted to survive diverse climatological and habitat conditions and requirements, especially in areas affected by humans (e.g. protected areas).
Despite the uneven spatial exploration in Croatia, we have compiled much information regarding invasive alien plants which threat national and European biodiversity. Although we are still working on this issue, we have decided to present our current knowledge about distribution patterns of invasive alien plants in Croatia, and thereby draw an attention to the invasive plants problem in this, till now “gap”, but almost central, area in the SE Europe. Our results indicated some important remarks, which should be taken into consideration not only on local, but also on a broader scale:
- • at least one invasive species is reported in almost 50% of the area of Croatia and on average, most of the MTB¼ grid cells contained five invasive plants;
- • a higher concentration of invasive alien plants (> 30) was noted in the major urban centres located at the intersection of main continental transport corridors and seaports;
- • number of invasive plants is increasing in the SE direction and reflects positive correlation with temperature: they occurred in a relatively wide altitude range, but mostly no higher than 1100 m a.s.l.; the largest proportion of recorded introductions and naturalization in the last 20 years has occurred in the Mediterranean region of Croatia, especially on islands;
- • almost 75% of all sites with invasive plants are located within a few habitats with direct anthropogenic influence, the most invaded habitats are agricultural areas, artificial surfaces, and affected forests; the number of invasive plants increased with habitat diversity.
These critical facts point to the importance of further permanent monitoring and researches of invasive alien taxa in Croatia, especially in the continental and Mediterranean urban areas, to ensure more efficiency in future management and control projects on the both national and European levels.
Disclosure of interest
The authors declare that they have no conflicts of interest concerning this article.
Acknowledgements
The research was financially supported through a grant from the Ministry of Science, Education and Sports of the Republic of Croatia (project 119-1191193-1227). We are grateful to numerous colleagues for any kind of contribution to the underlying inventory of plant taxa in Croatia. We also thank the numerous Flora Croatica Database collaborators who worked for almost two decades on the digitalization and geocoding of the herbarium collections ZA, ZAHO and CNHM. The authors would like to thank anonymous reviewers for their useful comments on the manuscript.