

1 Introduction
As outlined in a recent publication [1], until the early 1980s the scorpion fauna of French Guiana did not particularly call the attention of experts, and the few publications devoted to this fauna were limited to an isolated description [2] or to monographic compilations [3,4]. The first framework dedicated to the scorpion fauna of French Guiana was the one by Lourenço [5], in which all the species known to that date were treated. Many subsequent publications followed on the scorpion fauna of French Guiana, including some dealing with soil species [6]; however, very few studies were dedicated to the French Guiana Massifs represented by Inselbergs (Figs. 1–2). For a general overview on Tepuys and Inselbergs as possible palaeoclimate refugia and/or endemic centres, the reader may refer to Lourenço [1].

Aerial view of French Guiana rainforest in the region of the Mitaraka Massif.
Photo© Xavier Desmier.

Aerial view of the Massif of Mitaraka.
Photo© Xavier Desmier
The description of a new species of Ananteris from the Haut Ouarimapan, in the extreme southwest of French Guiana [7], represented the first contribution to the faunas of Inselbergs in this department. Moreover, even if the studies on the French Guiana scorpion fauna are far from being complete, this region appears as one of the ‘hot-spots’ for biodiversity in South America. The degree of endemism for the scorpion species present in the region can overpass 70% [8,9].
During the ‘Our Planet Reviewed’ Guyane-2015 expedition to the Mitaraka Massif in French Guiana, several scorpions were collected, among which two interesting specimens belonging to a new species of Ananteris. The Mitaraka Massif is a site located near the borders of French Guiana, Brazil, and Suriname (Fig. 3). These very small specimens were obtained with the use of extraction methods, and represent the second species of Ananteris found in this massif. The description of this new species brings further evidence about the biogeographic patterns of distribution presented by most species of the genus Ananteris, which are highly endemic in most biogeographic realms of South America, including the Tepuys and Inselberg massifs.
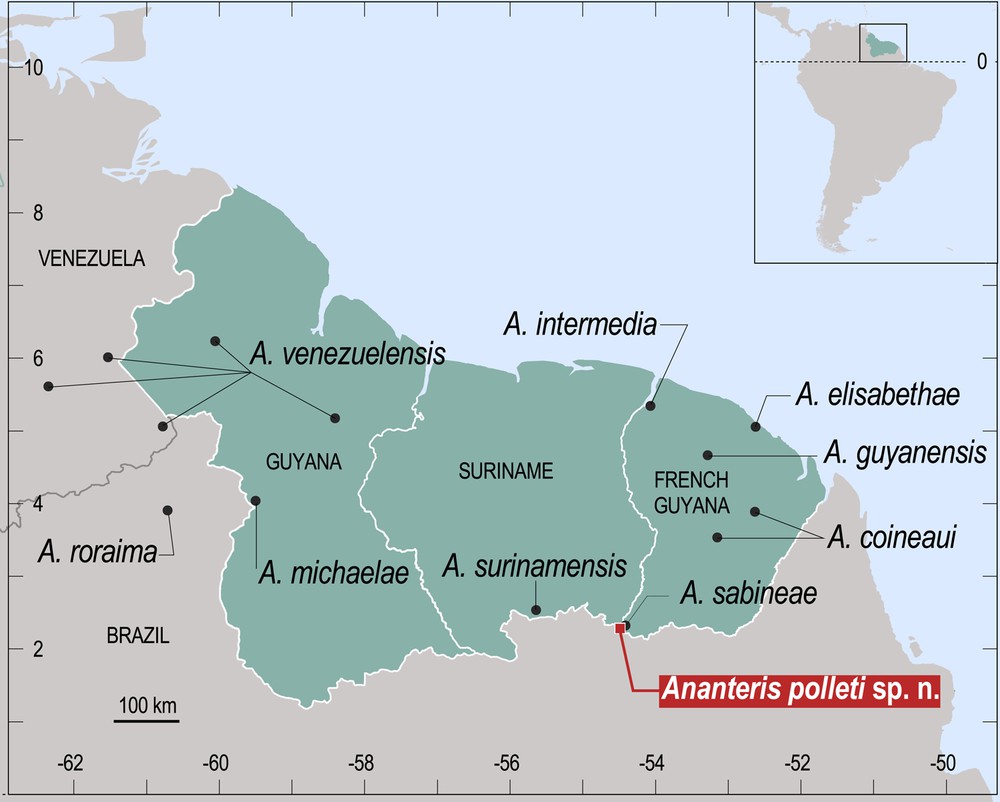
Map of the Guayana Floristic Province with the known distribution of the Ananteris species.
Map Lucienne Wilmé.
2 The genus Ananteris Thorell in the ‘Guayana Region’ and in French Guiana in particular
After the creation of the genus Ananteris Thorell, 1891 and the description of Ananteris balzanii Thorell, 1891, from the State of Mato Grosso in Brazil [10], the group remained inconspicuous by the number of species (three), and this until the 1970s. Nevertheless, the number of new species described within the genus steadily increased after its revision by Lourenço [11].
In recent years, only a rather limited number of species were described from the ‘Guayana Region’ [12]. Besides, most of the described species were from French Guiana [5,7,11,13,14], with a few exceptions from Brazil, Guyana, and Suriname [15–17]. The pace of descriptions from the Guyana region was much less intense than in other regions, mainly because the inventory work with the area is practically non-existent.
The species previously described from French Guiana are: Ananteris coineaui Lourenço, 1982, Ananteris guyanensis Lourenço & Monod, 1999, Ananteris sabineae Lourenço, 2001, Ananteris elisabethae Lourenço, 2003 and Ananteris intermedia Lourenço, 2012 [6,7,11,13,14]. Other species also described from the ‘Guayana Region’ are Ananteris roraima Lourenço & Duhem, 2010 from the Brazilian State of Roraima [17], Ananteris surinamensis Lourenço, 2012 from Suriname, Ananteris michaelae Lourenço, 2013 from Guyana and Ananteris venezuelensis González-Sponga, 1972 from Venezuela and Guyana [15,16,18] (Fig. 3). The description of a new species brings the total number of known Ananteris species described from the ‘Guayana Region’ to ten.
3 Methods
The scorpions were collected in the moist rainforest (Fig. 4) with the use of white water traps which are dug slightly into the soil or litter layer so that their upper rim reaches the surface (Fig. 5). The fixative used in site 1 was formaline 10% with detergent; in site 2, because of severe rainfall, salty water with detergent was used. Traps were serviced by pouring the yields through a small-mesh plastic sieve and gently transferring them (with a painter's brush) into a whirlpack and adding a 90% alcohol solution. In the lab, all invertebrate specimens were extracted in the usual way (M. Pollet, in litt.).

Moist rainforest, habitat of Ananteris polleti sp. n.
Photo© Marc Pollet.

Collection of samples from ‘white water traps’ in the field.
Photo© Xavier Desmier.
Illustrations and measurements were produced using a Wild M5 stereo-microscope with a drawing tube and an ocular micrometer. The measurements follow Stahnke [19] and are given in mm. Trichobothrial notations follow Vachon [20] and morphological terminology mostly follows Vachon [21] and Hjelle [22].
4 Taxonomic treatment
Family Buthidae C. L. Koch, 1837
Genus Ananteris Thorell, 1891
Ananteris polleti sp. n. (Figs. 6–8)

Ananteris polleti sp. n. Habitus of the male holotype. A–B. Dorsal and ventral aspects.

Ananteris polleti sp. n. Male holotype. A–B. Latero-dorsal and latero-ventral aspects. C. Metasoma, lateral aspect. D. Ventral aspect showing coxapophysis, sternum, genital operculum and pectines.
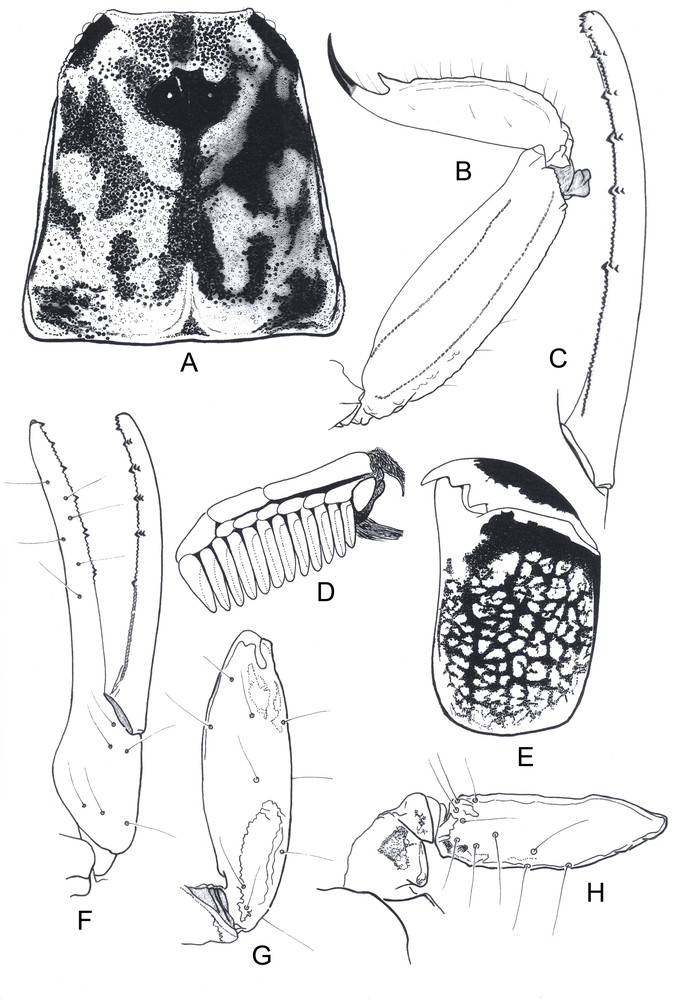
Ananteris polleti sp. n. Male holotype. A. Carapace. B. Metasomal segment V and telson, lateral aspect. C. Cutting edge of movable finger, showing rows of granulations. D. Pecten. E. Chelicera showing characteristic variegated pigmentation. F–H. Trichobothrial pattern. F. Chela, dorso-external aspect. G. Patella, dorsal aspect. H. Femur, dorsal aspect.
Type material: French Guiana, Mitaraka, ‘La Planète revisitée–MNHN/PNI Guyane 2015 (APA 973-1)’, (02°13’59.1” N, 54°26’37.9” W), 433 m, tropical moist forest–in plateau, 2/III//2015-8/III/2015 (M. Pollet), male holotype; (02°14’17.8” N, 54°27’08.2” W), 352 m, tropical moist forest–in slope, 25/II/2015-3/III/2015, (M. Pollet), male paratype. Material deposited in the ‘Muséum national d’histoire naturelle’, Paris.
Etymology: The specific name honours Marc Pollet (Research Institute for Nature and Forest, INBO, Brussels) who collected the type material.
Diagnosis: Species of very small size compared to the average size of the other species within the genus (14.66 mm in total length for male holotype; see morphometric values after the description). General coloration yellow to brownish–yellow, intensely marked with dark brown variegated spots. Pedipalps moderately short; fingers with six rows of granules; holotype and paratype pectines small with 11–11 and 12–12 teeth, respectively. Carinae and granulation moderately to strongly marked; dorsal and latero-dorsal carinae on metasomal segments II to IV with 3–4 posterior spinoids granules. Trichobothriotaxy, type A-β-beta.
Relationships: mainly by its diminutive size, aspects of the pigmentation pattern, and small pectines with a reduced number of teeth, the new species shows affinities with Ananteris festae Borelli, 1899 and Ananteris cisandinus Lourenço, 2015. These two species are respectively known only from Ecuador and the Amazonian/cis-Andean region of Peru. From the other species of the Guyana region, the new species can be distinguished by a combination of characters: (i) very diminutive size, (ii) small pectines with only 11 to 12 teeth in males, (iii) dorsal and latero-dorsal carinae on metasomal segments II to IV with 3–4 posterior spinoid granules. It differs from the only other Ananteris species also known from the Mitaraka Massif, A. sabineae, by the presence of an intense variegated pigmentation on the chelicerae (absent from A. sabineae), and by only 11–12 teeth on the pectines vs. 20 in A. sabineae.
Description based on the male holotype and the male paratype (morphometric measurements after the description).
Coloration. Generally yellow to brownish–yellow with brown to dark brown variegated pigmented zones on the carapace, the tergites, and the appendages. Prosoma: carapace yellow with dark brown spots on anterior, lateral and posterior edges; eyes surrounded by black pigment. Mesosoma yellow with confluent brownish zones on posterior and lateral edges of tergites. Metasomal segments I to V yellow; all segments marked with diffused brown spots. Vesicle yellow marbled with light brown zones; base of aculeus yellow, tip reddish. Venter yellow to pale yellow; coxapophysis and sternites infuscate. Chelicerae yellow with variegated blackish spots over the entire surface; fingers with blackish spots; teeth yellow. Pedipalps yellow; femur, patella, and chela strongly marked with dark brown spots; chela hand and fingers dark brown. Legs yellow, intensely marked with dark brown spots.
Morphology. Carapace with moderately to strongly marked granulation; anterior margin almost straight. Anterior median superciliary and posterior median carinae weak or absent. All furrows moderate to weak. Median ocular tubercle distinctly anterior to the centre of carapace; median eyes separated by approximately one ocular diameter. Three pairs of reduced lateral eyes. Sternum subpentagonal. Mesosoma: tergites with moderately to strongly marked granulation, similar to that of carapace. Median carina moderately to weakly marked on all tergites. Tergite VII pentacarinate. Venter: genital operculum divided longitudinally, each plate more or less suboval in shape. Pectines small: pectinal tooth count 11–11 in male holotype (12–12 in male paratype); basal middle lamellae of pectines not dilated; fulcra absent. Sternites smooth; only VII slightly granular; spiracles rather short; setation moderate; sternite VII with very weakly marked carinae and granulation. Metasomal segments I to III with 10 carinae, crenulate; segment IV with 8 carinae, crenulate; segment V slightly rounded and smooth, with vestigial carinae; intercarinal spaces moderately granular on all segments; dorsal and latero-dorsal carinae on segments II to IV with 3–4 posterior spinoid granules. Telson elongate and smooth; aculeus short and weakly curved; subaculear tooth strongly marked and spinoid. Cheliceral dentition characteristic of family Buthidae [23]; fixed finger with two strong basal teeth; movable finger with two vestigial basal teeth; ventral surface of both finger and manus with long, dense setae. Pedipalps: femur pentacarinate; patella and chela with weak to vestigial carinae; internal face of patella with only vestigial spinoid granules; all faces weakly granular, almost smooth. Fixed and movable fingers with 6, almost linear, rows of granules; two small external and one internal accessory granule present at the base of each row; three granules at the extremity of the fingers. Trichobothriotaxy; orthobothriotaxy A-β-beta [20,24]. Legs: Tarsus with very numerous, fine, median setae ventrally. Tibial spurs strongly developed on legs III and IV.
Morphometric values (in mm) of the male holotype. Total length including telson, 14.66. Carapace: length, 1.87; anterior width, 1.13; posterior width, 1.73. Mesosoma length, 3.47. Metasomal segments. I: length, 0.93; width, 1.07; II: length, 1.00; width, 1.10; III: length, 1.13; width, 1.13; IV: length, 1.33; width, 1.13; V: length, 2.53; width, 1.27; depth, 0.93. Telson length, 2.40; width, 0.73; depth, 0.67. Pedipalp: femur length, 1.63, width, 0.53; patella length, 1.87, width, 0.60; chela length, 2.47, width, 0.43, depth, 0.50; movable finger length, 1.80.
Disclosure of interest
The author declares that he has no competing interest.
Acknowledgements
The material related to the new species was collected during the ‘Our Planet reviewed’ Guyane-2015 expedition in the Mitaraka range, in the core area of the French Guiana Amazonian Park, organized by the MNHN and Pro-Natura international. The expedition was funded by the European Regional Development Fund (ERDF), the Conseil régional de Guyane, the Conseil général de Guyane, the Direction de l’Environnement, de l’Aménagement et du Logement and by the Ministère de l’Éducation nationale, de l’Enseignement supérieur et de la Recherche. It was carried out in collaboration with the ‘Parc amazonien de Guyane’ and the ‘Société entomologique Antilles–Guyane’. My thanks are also due to Xavier Desmier for permission to use his photos of the Mitaraka Massif (Figs. 1, 2, 5), to Lucienne Wilmé (Missouri Botanical Garden, Madagascar) for the preparation of the map (Fig. 3), to Marc Pollet, for permission to use one of his photos of the habitat (Fig. 4) and information on the collecting methods, and finally to Élise-Anne Leguin, MNHN (Paris) for the preparation of the photos and plates of the new species.