

1 Introduction
Quadrupeds use two kinds of gaits [1]. Symmetrical gaits (e.g., pace, walk or trot) occur when footfalls of two feet of the fore pair and of the hind pair are evenly spaced in time. Asymmetrical gaits (e.g., gallop and bound) occur when the actions in at least one pair are unevenly spaced in time [2]. Classical methods allowed the comparison of symmetrical gaits [2–4], but there was only one method available to describe all asymmetrical gaits [5]. Furthermore, most of the parameters used for symmetrical and asymmetrical gait analyses were different thus there was no way of analysing all the gaits using the same method. All methods were based on the stride, that is the cycle of one reference foot beginning with the footfall (or any given action) of this foot and ending with the next footfall of the same foot. The actions of the three other feet had to begin during that stride [6]. It was sometimes difficult to establish which foot must be chosen as reference, and depending on the study this needed be a hind [2,4–11] or forefoot [12]. It was furthermore not always possible to establish a reference foot for a whole study, or even for a complete sequence if the animal changed its gait [13]. Thus there existed no method able to encompass the observed phenomena as a whole.
2 Method
A new approach is proposed, which is based on two observations. First, in all vertebrates, due to bilateral symmetry, there is a total morphological and functional similarity between both limbs of a pair, fore or hind. Alternatively, fore limbs and hind limbs can be very different morphologically and functionally, in bats for example. The strong morphological relationship within pairs means that they must be linked by a basic coordination and thus two limbs of a pair must not be separated in the analysis. Secondly, locomotion is initiated by an activity of the brainstem locomotor system, which activates a locomotor pattern generator in the spinal cord [1]. Thus, for locomotor initiation, there is rostro-caudal activation, which leads to the forelimb spinal network being activated before the hind limb spinal network. Furthermore, in terms of space, the fore limbs are the first to approach modifications of the environment and must be the first to adjust their movements. The analysis must take into account these two factors and describe the whole locomotor sequence, beginning by the action of the fore limbs, followed by the action of the hind limbs. In classical methods, the analysis was based on the stride, which does not include the whole sequence, and thus the stride often contained the action of a limb that does not belong to the sequence of the reference foot. The cyclic interpretation of the feet coordination in classical methods involves a dissociation of the coordination of paired feet and a caudo-rostral coordination (Fig. 1).
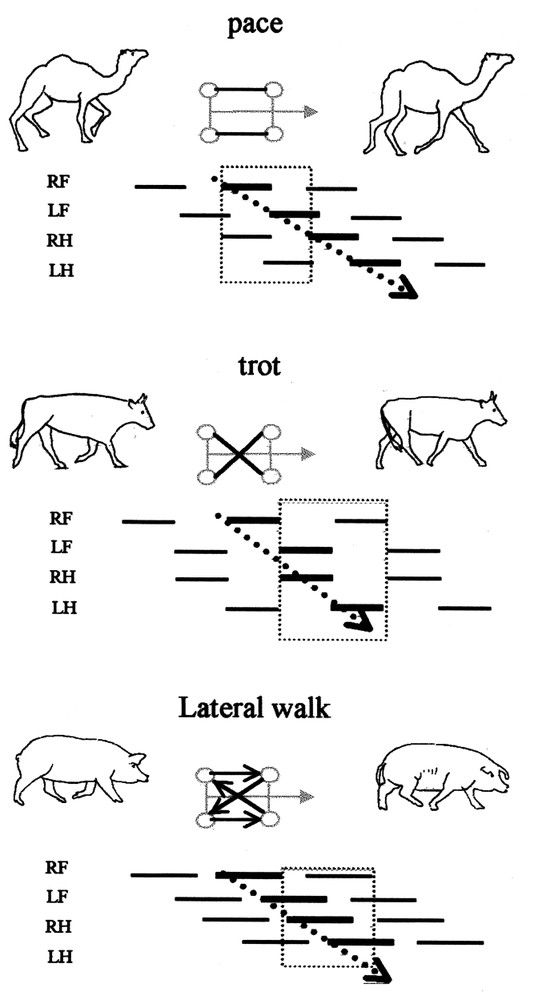
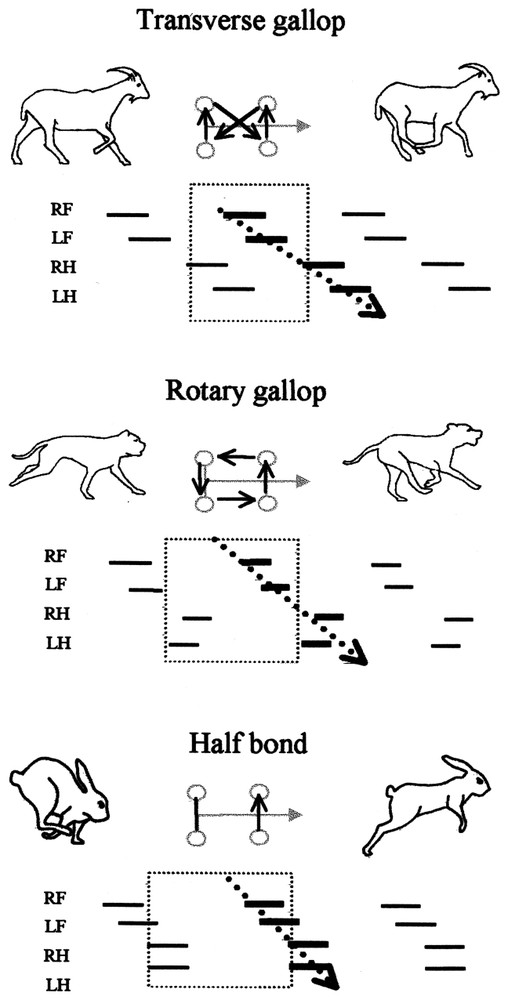
Differences between the antero-posterior sequence analysis and the stride analysis in gaits. Gaits are represented on gait diagrams [18]. The four feet are plotted on successive lines, the time when a foot is on the ground is symbolised by a continuous line. RF: First fore foot (arbitrary right); LF: second fore foot (left); RH: first hind foot (right); LH: second hind foot (left). On the schemas between animal silhouettes, the grey arrow represents the head, the circles represent the feet. The black lines linking the circles represent simultaneous footfalls and the black arrows the order of succession of the footfalls during a stride. The use of the stride (included in the dotted square limits on the gait diagrams) gives the succession of the footfalls presented in the scheme. It highlights the differences between the gaits, and suggests diagonal and postero-anterior coordination. On the contrary, the use of the antero-posterior sequence (thick lines on the gait diagrams) using the movements of the two fore feet followed by the movements of the two hind feet highlights the same basic coordination pattern for all the gaits (dotted arrow).
The use of six variables in the antero-posterior sequence, five of which are already used by Hildebrand to described symmetrical gaits [2], enables the representation of all gaits, symmetrical and asymmetrical [14]. Three of the five are time lags, and include that (i) between the forefeet footfall (F lag), (ii) between the hind feet footfall (H lag), and (iii) between the footfall of ipsilateral feet, fore and hind feet of the same side (P lag). The velocity is expressed by the time a foot is on the ground, the duty factor, which decreases when speed increases [15]. The duty factor is similar for the four feet in symmetrical gaits [2], but not in asymmetrical gaits [8,12,16] and thus (iv) the fore limbs' duty factor, and (v) the hind limbs' duty factor must be considered separately. All measurements can be expressed as a percentage of (vi) the cycle duration, which is the same for all the feet in a sequence.
3 Interpretation
The graphical representation of gaits using the antero-posterior sequence allows a new interpretation of the motor coordination (Fig. 2). All gaits follow the antero-posterior sequence perfectly, with fore feet action followed by that of the hind feet. All the gaits can be identified using three parameters: the F lag, the H lag and the P lag (Table 1).
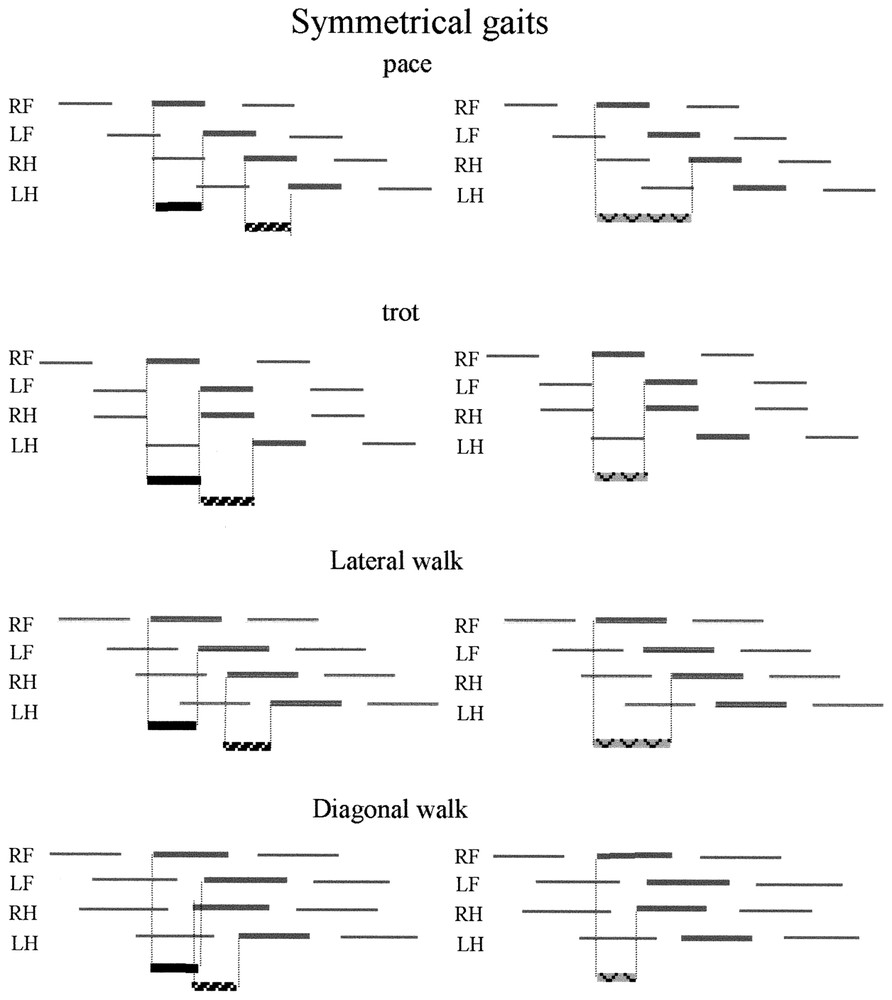
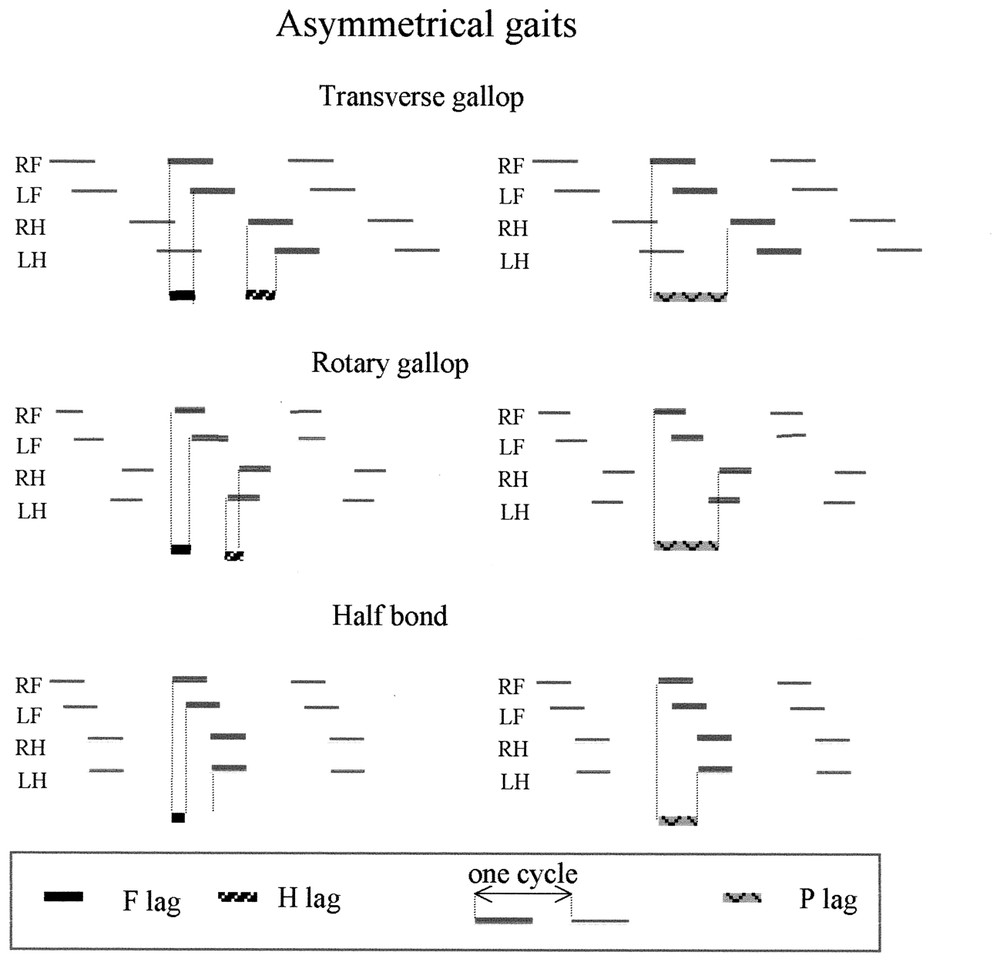
Characterisation of the gaits using the antero-posterior sequence. Inside the basic coordination pattern, the gaits are generated by adjustments of the time lag between the fore feet touch down (black line: F lag), between the hind feet touch down (hatched line, H lag) and between the pairs touch down (pointed black line: P lag). (See data in Table 1.) In symmetrical gaits, by definition [2] F lag = H lag = 50%. The different gaits arise from difference in the time lag between the pair movements. P lag = 100% produces a pace: the movement of the right hind foot is synchronized with the movement of the right fore foot of the next sequence. P lag = 50% produces a trot and the simultaneous movements of diagonal feet (fore and controlateral hind) arises from the synchronisation of movements of the feet inside the sequence (LF and RH) and from the coupling of successive sequences (LH of a sequence with the RF of the precedent sequence). P lag >50% produces a lateral walk, whereas P lag <50% produces a diagonal walk. In asymmetrical gaits, the tendency to synchronise paired feet [1] is expressed by a decrease of the fore and hind lags (<50% cycle duration). Those time lags (F lag and H lag) are used to characterize the gaits (Table 1), whereas the time lags between the movements of the two pairs (P lag) depend on speed. H lag in rotary gallop is negative, because the first hind foot (RH) touches the ground after the second hind foot (LH).
Definition of the gaits using the antero-posterior sequence. The gaits can be defined by the time lag between the footfalls of the forefeet (F lag), the footfall of the hind feet (H lag), and between the footfall of the first fore foot and the first hind foot (P lag). The time lag can be expressed as a percentage of the cycle duration, which is the same for the four feet. (See graphical illustration in Fig. 2)
| Gait | F lag | H lag | P lag |
| Pace | 50 | 50 | 100 |
| Trot | 50 | 50 | 50 |
| Lateral walk | 50 | 50 | >50 |
| Diagonal walk | 50 | 50 | <50 |
| Transverse gallop | <50 | <50 | depends on the speed |
| Rotary gallop | <50 | <0 | depends on the speed |
| Half bond | <50 | 0 | depends on the speed |
| Bond | 0 | 0 | depends on the speed |
| Pronk | 0 | 0 | 0 |
A gait variability of 5% around the given values has been established [2]. For symmetrical gaits, by definition [2] the time lags between the two feet of the pairs (F lag and H lag) are the same (50% of the cycle duration). The succession of the movements, simultaneous of the ipsilateral feet in the pace, simultaneous of the diagonal in the trot and the alternated in the walks [17] are the result of the time lag between the action of the fore and hind pairs (P lag). The P lag in pace (100% cycle duration) induces simultaneous movements of the two right feet and of the two left feet. The forefeet are synchronized with the hind feet of the precedent sequence. In the trot, the P lag (50% cycle duration) induces a simultaneous action of the diagonal feet; i.e. second foot of a pair and first foot of the other pair. The second forefoot is synchronized with the first hind foot in a sequence, whereas the first fore foot is synchronized with the second hind foot of the precedent sequence. During walking, alternated movements of the limbs arise from an intermediate time lag; i.e. more than 45% in a lateral walk, less than 45% in a diagonal walk [2]. Once again, the coupling of successive sequences induces a temporal superposition of the limb movements, mainly diagonally, that provides good stability of the animal during its locomotion [14].
During asymmetrical gaits, animals modify more parameters, including the pair lags as in symmetrical gaits, but also coordination between the two fore feet and between the two hind feet. This coordination changes from the paired feet strictly alternated movement in symmetrical gaits (pair lag = 50%) to an in phase coordination of paired feet in asymmetrical gaits, which is completed in bounds [1]. Thus, the F lag and the H lag are lower than 50% of the cycle and are used to determine the gait (Table 1). During asymmetrical gaits, both pairs are not always coordinated in the same way, and in the half bound, the hind feet are synchronized, whereas the movements of the fore feet are not simultaneous. The P lag varied with speed during asymmetrical gaits. The tendency to synchronize the paired feet associated with the decrease of the duty factor allows the introduction of suspension, during which no foot is placed on the ground. Depending on speed and animal morphology, suspension can occur after the stance of the fore feet (gathered suspension), after that of the hind feet (extended suspension), or with both [14]. The rotary gallop is different from all other gaits in that the first foot to touch the ground is the right fore, while the first hind is a left foot. It thus appears that the hind feet are ‘left-handed’, while the forefeet are ‘right-handed’ and vice versa.
4 Conclusion
The use of the stride for gait analysis allowed classifying them by highlighting their differences. The stride hides the common pattern, which becomes conspicuous when using the antero-posterior sequence. Thus, leaving the stride paradigm through the use of the antero-posterior sequence provides a simple method to analyse all gaits coordination, symmetrical as well as asymmetrical and open a new framework for locomotion analysis.
Acknowledgements
I wish to thank P.Y. Gagnier, M. Herbin, J.-P. Gasc and S. Renous for their comments and advices. Special thank to T. Sharbel for his generous help.