

Version française abrégée
1 Introduction
Comme on l'a déjà exposé dans une publication récente, les scorpions humicoles sont globalement rares. Parmi les Buthoidea ont été signalés les cas d'Akentrobuthus leleupi Lamoral, espèce endémique des forêts de la province du Kivu au Congo, mais aussi celui de Lychasioides amieti Vachon, décrite de la forêt d'Otomoto au Cameroun. Dans cette même publication, un nouveau genre et une nouvelle espèce de micro-scorpion Buthidae humicole, Microananteris minor Lourenço, ont été décrits de la Guyane française, représentant ainsi le premier élément humicole connu pour la région néotropicale.
Parmi les micro-Buthoidea récemment décrits de Madagascar, deux nouveaux genres, Microcharmus Lourenço et Neoprotobuthus Lourenço, sont composés uniquement de vraies espèces humicoles. La découverte de plusieurs espèces nouvelles de Microcharmus a permis d'isoler ce groupe de micro-scorpions dans une famille à part, celle des Microcharmidae Lourenço. Un troisième genre malgache de micro-Buthoidea, Pseudouroplectes Lourenço, a été placé dans la famille des Buthidae. À présent, un nouveau genre et une nouvelle espèce de micro-scorpion Microcharmidae humicole, Ankaranocharmus pauliani gen. n., sp. n., sont décrits du Nord de Madagascar. L'analyse comparative des papilles sensorielles des peignes confirme la position du nouveau genre chez les Microcharmidae, mais aussi celle des Pseudouroplectes chez les Buthidae.
2 Analyse comparative des papilles sensorielles des peignes
Ainsi que j'ai pu l'expliquer dans ma précédente note sur les scorpions humicoles, la majorité des études existantes sur les papilles sensorielles des peignes de scorpions font référence à leur rôle de chémorécepteur. Cependant, les particularités morphologiques des papilles sensorielles ont également une valeur dans des études de phylogénie ou d'adaptation écologique.
Les observations faites pour différentes espèces de Buthidae malgaches, comme celles du genre Grosphus, montrent des papilles avec une forme de spatule, en général assez courtes (Figs. 1–4). Chez le genre Pseudouroplectes, les papilles ont toujours une forme de spatule, bien qu'elles soient un peu plus allongées (Figs. 5–8). L'observation des papilles sensorielles de deux espèces humicoles appartenant à la famille des Microcharmidae, Neoprotobuthus intermedius (Figs. 9–12) et Microcharmus fisheri (voir Figs. 13–16 de ma précédente note), montrent une structure arrondie, en forme de bouteille. Chez les nouveaux genre et espèce décrits à présent, les papilles ont une forme également arrondie, mais le profil de bouteille est moins marqué, et leur taille globale est plus petite que celles observées pour M. fisheri et N. intermedius (Figs. 12 et 16). La présente étude à l'aide du microscope électronique à balayage semble confirmer le modèle déjà observé pour d'autres espèces humicoles.




Grosphus olgae Lourenço (female paratype): (1), pecten; (2–4), microstructure of peg-shaped sensillae on teeth.
Grosphus olgae Lourenço (femelle paratype) : (1), peigne ; (2–4), microstructure des papilles sensorielles des dents.

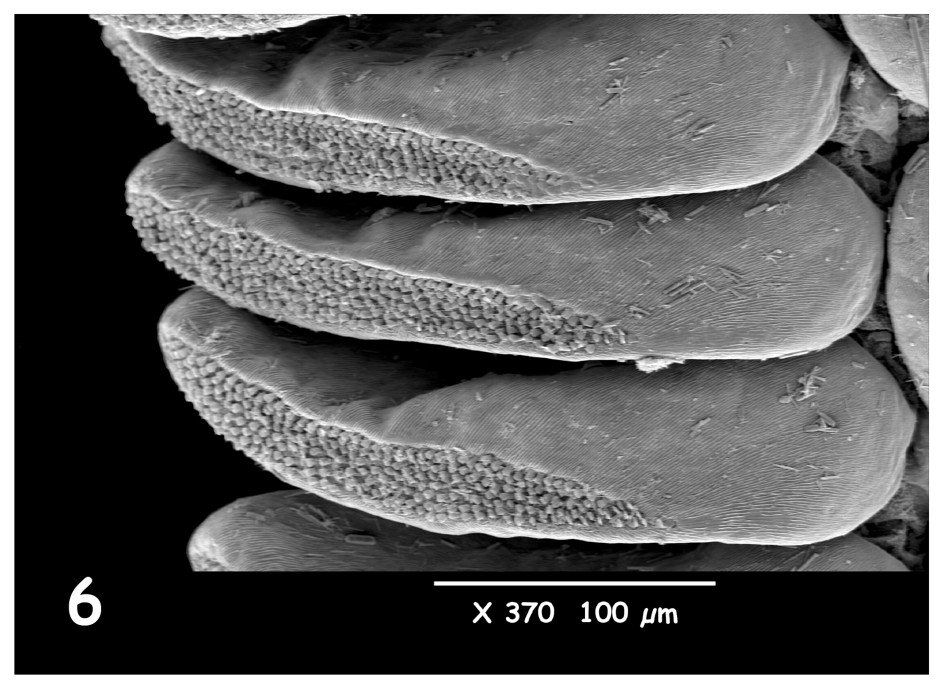


Pseudouroplectes betschi Lourenço (femelle paratype). Idem Figs. 1–4.
Pseudouroplectes betschi Lourenço (female paratype). Idem Figs. 1–4.




Neoprotobuthus intermedius Lourenço (female paratype). Idem Figs. 1–4.
Neoprotobuthus intermedius Lourenço (femelle paratype). Idem Figs. 1–4.




Ankaranocharmus pauliani gen. n., sp. n. (female holotype). Idem Figs. 1–4.
Ankaranocharmus pauliani gen. n., sp. n. (femelle holotype). Idem Figs. 1–4.
1 Introduction
As pointed out in a recent publication [1], humicolous scorpions are globally rare. A few genuine cases have been reported among buthoid scorpions. These include Akentrobuthus leleupi Lamoral, an endemic species from the forests of the Kivu Province in Congo and Lychasioides amieti Vachon from the forest of Otomoto, Cameroon. In my previous paper [1], a new genus and species of humicolous micro-buthid scorpion, Microananteris minor Lourenço, was described from French Guyana. This represents the first humicolous element to be recorded from the Neotropical region.
Among the micro-buthoids recently described from Madagascar, two new genera Microcharmus Lourenço and Neoprotobuthus Lourenço are composed uniquely of genuine humicolous species. With the discovery of several new species of the genus Microcharmus, this group of micro-scorpions has been accommodated in its own family, Microcharmidae Lourenço [2–6]. A third micro-buthoid genus from Madagascar, Pseudouroplectes Lourenço, was accommodated in the family Buthidae [2,3]. In the present paper, a new genus and species of humicolous microcharmid scorpion, Ankaranocharmus pauliani gen. n., sp. n., is described from the North of Madagascar. Comparative analysis of the microstructure of the peg-sensillae of the pectines by scanning electron microscopy, confirms the position of the new genus within the family Microcharmidae. It also clarifies the position of Pseudouroplectes in the buthids.
2 Comparative analysis of microstructure of the peg-shaped sensillae of the pectines in some groups of buthoid scorpions
As I have already explained [1], most existing studies of the peg-shaped sensillae of scorpion pectines are concerned with their mechano- and chemosensory roles [7]. However, their structure can also be useful in studies of phylogeny and adaptation. Several species in different genera and families have already been investigated by scanning electron microscopy. Observations carried out on different species of Malagasy buthids, such as those of the genus Grosphus (Figs. 1–4), reveal spatula-like peg-shaped sensillae, which are generally very short. In the genus Pseudouroplectes the peg-shaped sensillae are also spatula-like, but they are slightly longer than those of Grosphus (Figs. 5–8). The peg-shaped sensillae of two humicolous species of the family Microcharmidae, Neoprotobuthus intermedius (Figs. 9–12) and Microcharmus fisheri (see Figs. 13–16 in [1]), have a rounded structure, more like the shape of a bottle.
In Ankaranocharmus pauliani gen. n., sp. n., described here, the peg-shaped sensillae are equally rounded, but the bottle-like shape is less marked and their overall size is smaller that those of M. fisheri and N. intermedius (Figs. 12 and 16). The present study of peg-shaped sensillae by means of SEM seems to confirm the pattern already observed in others humicolous species.
3 Taxonomic treatment
Ankaranocharmus gen. n.
Diagnosis. Very small scorpions, when compared with the average size of most micro-buthoid genera, and measuring 14.7 mm in total length (see morphometric values). Sternum pentagonal; spiracles short but semi-slit-like; distal tooth of pectines less rounded than in other microcharmids; fulcra absent; tibial spurs moderate on leg IV, but absent from leg III; telson small and long, without subaculear tooth; trichobothriotaxy, A-α (alpha), orthobothriotaxic.
Type species of the genus. Ankaranocharmus pauliani sp. n. (Figs. 13–19).
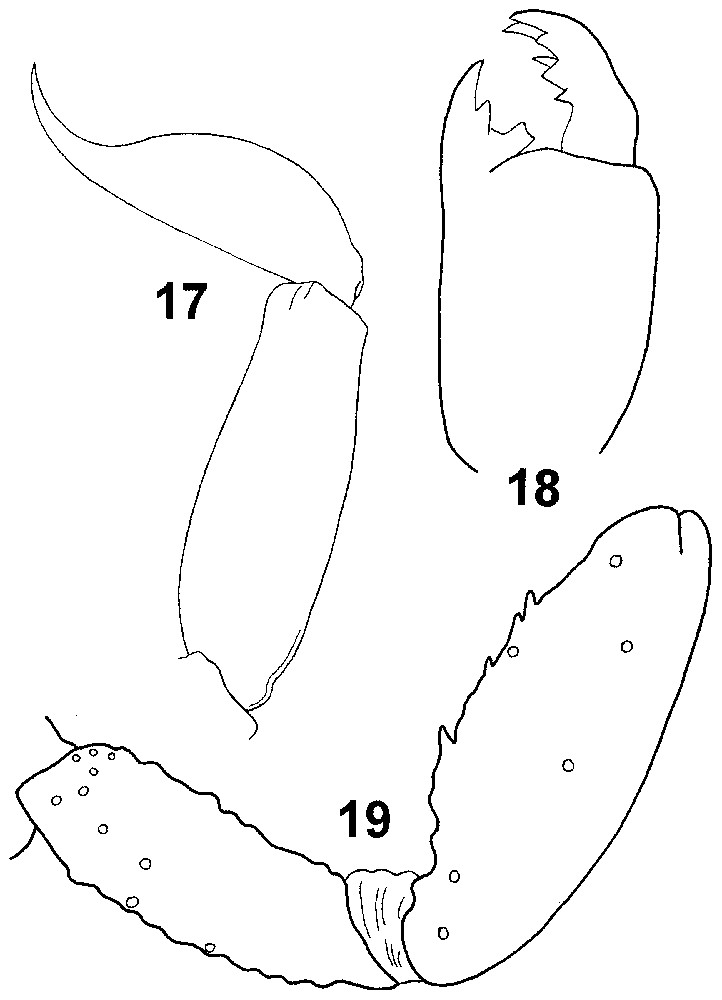
Ankaranocharmus pauliani gen. n., sp. n. (female holotype). (17) Metasomal segment V and telson, lateral aspect; (18) chelicera, dorsal aspect; (19) femur, and patella dorsal aspect, showing alpha configuration for dorsal trichobothria of femur.
Ankaranocharmus pauliani gen. n., sp. n. (holotype femelle). (17) Ve anneau du metasoma et telson, vue latérale ; (18) chélicère, vue dorsale ; (19) fémur et tibia en vue dorsale, avec les trichobothries dorsales du fémur en configuration alpha.
Type-material female holotype. Madagascar, ‘Pro-vince d'Antsiranana’, ‘Réserve spéciale d'Ankarana’, ‘Encampement des Anglais’ (Anilotra), IX/2001 (W.R. Lourenço leg.). Undisturbed Tsingy forest. In litter. No paratypes. Deposited in the ‘Muséum national d'histoire naturelle’, Paris.
Etymology. The specific name is in honour of Prof. Renaud Paulian, for the important contribution he has made to the zoology of Madagascar. Prof. Paulian has also assisted other national and foreign researchers and played a paramount role in the advancement of young French and Malagasy field scientists.
For data on the ecology and biogeography of the Ankarana Massif, refer to Lourenço and Goodman [8].
Description. Coloration. Basically brownish-yellow, symmetrically marbled with dark brownish markings, producing an overall spotted appearance. Prosoma: carapace yellowish, almost totally covered with brown spots; eyes surrounded by black pigment. Mesosoma: yellowish-brown with four longitudinal brown stripes. Metasoma: segments I to V yellowish, with several brown spots; segment V with more marked spots. Telson: vesicle yellowish without spots; aculeus reddish. Venter yellowish, with diffuse brown spots over the coxapophysis, sternum, genital operculum, pectines and sternites III to VII. Chelicerae yellowish with variegated spots over their entire surface; more pronounced anteriorly; fingers yellowish brown with reddish teeth. Pedipalps: yellowish densely marked with brownish spots, especially on the femur and patella; chela yellowish; fingers brownish with the rows of granules slightly reddish. Legs yellowish, densely marked with brownish spots.
Morphology. Carapace with an intense but thin granulation; anterior margin with a median concavity. All carinae and furrows weak. Median ocular tubercle distinctly anterior to the centre of carapace; median eyes separated by approximately one ocular diameter. Three pairs of lateral eyes. Sternum pentagonal. Mesosoma: tergites with a thin granulation. Median carina moderate to weak in all tergites. Tergite VII pentacarinate. Venter: genital operculum divided longitudinally, each plate having a more or less subtriangular shape. Pectines very small: pectinal tooth count 11–11; basal middle lamellae of the pectines not dilated; fulcra absent. Sternites smooth with short semi-slit-like spiracles; VII with a few granulations and vestigial carinae. Metasoma: segments I to III with 10 carinae, crenulate. Segment IV with 8 carinae, crenulate. Intercarinal spaces weakly granular. Segment V with five carinae. Telson with a ‘pear-like’ shape, almost smooth, without carinae, covered with long setae; aculeus short and moderately curved; subaculear tooth absent. Cheliceral dentition more or less characteristic of the buthoids [9]; fixed finger with two moderate basal teeth; movable finger with one weak basal tooth; ventral aspect of both finger and manus with dense, long setae. Pedipalps: femur pentacarinate; patella and chela with a few vestigial carinae; internal face of patella with some spinoid granules, two of which are strong; all faces weakly granular, almost smooth. Fixed and movable fingers with 7/8 almost linear rows of granules; two accessory granules present at the base of each row; extremity of movable fingers with 2 accessory granules. Trichobothriotaxy; orthobothriotaxy A-α(alpha) [10,11]. Legs: tarsus with very numerous fine median setae ventrally. Tibial spurs moderate on leg IV, absent on leg III.
Morphometric values (in mm) of the described new species. Total length, 14.7. Carapace: length, 1.94; anterior width, 1.13; posterior width, 2.00. Metasomal segment I: length, 1.00; width, 1.13. Metasomal segment V: length, 2.31; width, 0.88; depth, 0.81. Vesicle: width, 0.63; depth, 0.63. Pedipalp: femur length, 1.56, width, 0.50; patella length, 2.06, width, 0.69; chela length, 2.75, width, 0.56, depth, 0.56; movable finger length, 1.81.
Acknowledgements
I am very grateful to Laurent Albenga and Régis Cleva, ‘Département de systématique et évolution’, ‘Muséum national d'histoire naturelle’, Paris, for their help in the preparation of the SEM illustrations, and to Profs. John L. Cloudsley-Thompson, London, and J.-M. Betsch, ‘Muséum national d'histoire naturelle’, Paris, for reviewing the manuscript.