

1 Introduction
Through the impetus of Professors Doumandi and Doumandji-Mitiche from the Institut National Agronomique in El-Harrach (Algiers), acridian fauna (Orthoptera, Acridomorpha) have been the subject of various works for more than 20 years, due to the dramatic damage of these insects against crops in Algeria. In the North part of the country, we can quote [1–5]. In the oriental part of Algeria, several studies were devoted to the area of Biskra: [6–9]. As for Saharan oases, information is given in the following references: [10–13]. Our work proposes to compare the assemblages of grasshoppers within cultivated (oases) and steppic areas in the East of Algeria. Brague-Bouragba et al. [14] showed that in a pre-Saharan steppe in Algeria, reforestation did not affect biological diversity of coleopteran and spiders but dramatically changed the functioning of arthropod communities. The problem we address is to know if the fauna in anthropised environments results from the degradation of steppe fauna or if it exhibits its own particularities. In addition, it is interesting to compare the seasonal variations of assemblages between these two ecosystems, given that the steppe habitats undergo more the variations of rainfall than the anthropised areas, which are irrigated by man.
For this reason, it is useful to study several biological features of this fauna, in particular its phenology. The most common cycle of grasshoppers in the northern hemisphere consists in one annual generation, with a peak of adults observed in June-July [15], the wintering being ensured by diapausing embryos contained in eggs deposited in the ground [16,17]. In areas with milder winters, it is expected to find alternatives, such as for example the existence of several generations per annum, the egg being the form of resistance. Another possibility consists in a diapausing phase during winter ensured by instars or adults [18,5]. Our purpose is thus to get insights into the analysis of arid faunas by the comparison of phenology types present in the cultivated zones, irrigated or not, and in “natural” steppic zones.
Our study is centered on the area of Biskra, a locality at the altitude of 124 m A.S.L., situated at the North-East of the Algerian Sahara, between the Saharan Atlas in the North, the high plateaus and the Sahara in the South, at the limit between the arid and Saharan bioclimatic stages [9]. The natural environments of this area include diversified steppes, according to the presence of sand, emergent rock, various salt concentrations in soil, and also more or less salted wet depressions. In oases, the date palm cultivation is the traditional agriculture [19] and an original flora develops, often including halophytes favored by the salt increased by irrigation, and adventitious therophytes related to the cultures themselves [20].
2 Material and methods
In the area of Biskra, we made our studies in two types of environments, oasis (five stations) and steppe (two stations) (Table 1, Fig. 1). The stations (A1, A2, A3, C1 and C2) are situated at Zab El-Guebli (south of Zab Mountains). They are oases located on the southern part of the valley of Oued Djedi Wadi, where there is a current of water below sands. The stations (A1, A2 and A3) are close to Ben-Thious (East of Mekhadema) and the stations C1 and C2 near Mekhadema. The A1 station is an uncultivated surface of approximately 3 ha, located between two palm plantations. The vegetation has a non-degraded structure, characterized by a dense shrubby layer with the predominance of the Salsolaceae family. The dominant species are Salsola vermiculata, Salsola tetragona and Atriplex halimus. Mesembryanthemum nodiflorum is very abundant during the rainy season. Likewise, the A2 station is also an uncultivated surface of approximately 1 ha, previously exploited for the culture of barley, located in the area of Ben-Thious. Vegetation cover consists in shrubs, especially of Suaeda vera with a few individuals of S. vermiculata, A. halimus and Peganum harmala. As in the preceding station, M. nodiflorum is very frequent during the rainy season. The A3 station is an abandoned palm plantation, of surface of approximately 1 ha. The irrigation is occasional and in a random way. Intercalated surface between trees shows an almost bare ground, except near the seguias (irrigation channels), which are colonized by grasses. The presence of crusts saltworks is noticeable. Vegetation cover consists of several layers: the upper layer is represented by the date palm tree (Phoenix dactylifera), the shrubby layer by A. halimus, and the herbaceous one is dominated by Cynodon dactylon. The C1 station is a young palm plantation, of surface of approximately 4 ha, characterized by plantations separated by about 8 m. The irrigation is done regularly. The larger part of the intercalated surface is colonized by dense grasses among which, Poaceae are the most abundant family (wet biotope). But it should be noted that the vegetation is always under the influence of human activity, since it is an irrigated culture especially disturbed by the regular effect of weeding. There is no bush. The C2 station is a modern palm plantation in its first years of development, of surface of approximately 1 ha, with a drop-by-drop irrigation. In addition to the date palm trees, the flora is diversified with the predominance of Salsolaceae and Poaceae, particularly abundant near the trunks. The last stations are two bushy steppes grazed by cameline, ovine and caprine. They are located in the area of Bir-Naam, at Piedmont of the Saharan Atlas, 55 km west of the town of Biskra. The station Ca is a stony surface and its vegetation comprises a shrubby layer dominated by Astragalus armatus and a rich herbaceous layer, where Lygeum spartum, Farsetia occidentalis and Plantago albicans are the most abundant species. The station Sa is a sandy surface (stony ground covered with sand), where vegetation consists in tufts of Stipa tenacissima, with a high presence of Onopordum arenarium, Ononis serrata and Linaria aegyptiaca.
Summary of studied sites.
| Station | Habitat | Altitude (m) | Longitude | Latitude |
| A1 | OASIS | 76 | 5°29′55.26″E | 34°38′15.88″ N |
| A2 | OASIS | 76 | 5°29′53.60″E | 34°38′19.59″ N |
| A3 | OASIS | 76 | 5°29′57.67″E | 34°38′19.71″ N |
| C1 | OASIS | 76 | 5°28′22.27″E | 34°37′30.96″ N |
| C2 | OASIS | 76 | 5°28′23.51″E | 34°37′27.57″ N |
| CA | STEPPE | 222 | 5°8′34.45″ E | 34°45′6.72″ N |
| SA | STEPPE | 222 | 5°8′4.65″ E | 34°45′5.35″ N |
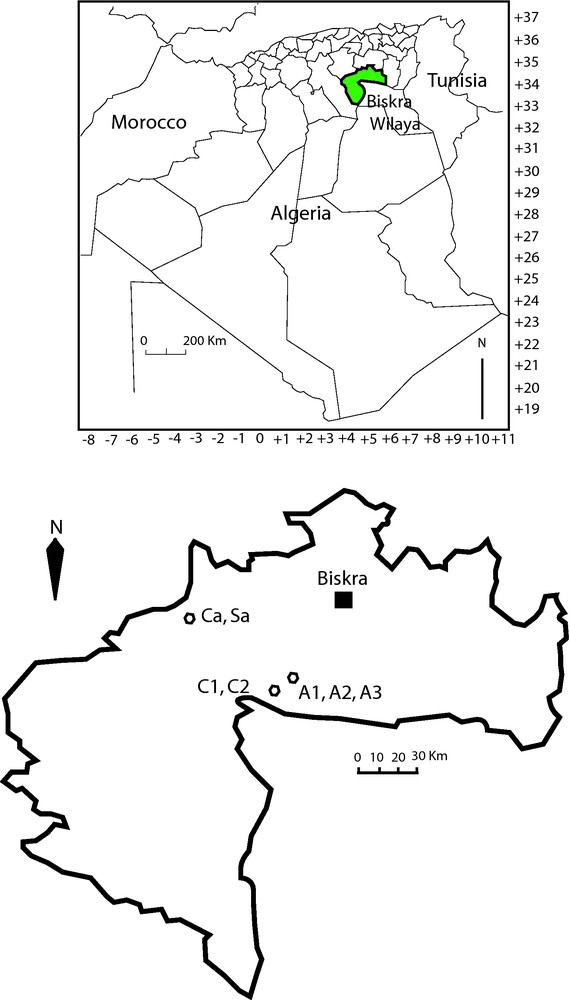
Map of the Wilaya of Biskra and position of study stations.
2.1 Climatology
In the area of Biskra, precipitations are weak and irregular for months and years. The rains are mainly observed in autumn and winter. The relative humidity of air varies appreciably according to the seasons. During summer, it falls up to 25% in July, resulting in a strong evaporation. On the other hand in winter, it rises up to 60%. The winds blow all year long, and the prevailing ones blow from the northwest. The south winds are generally cold and dry in winter. They are hot and very dry in the summer period (sirocco), increasing evaporation and thus dryness.
The climatic data were collected over the period 1985–2009 from the Office of Algerian meteorology. We used the ombrothermic diagram of Gaussen [21], which defines empirically a dry month when the total of the precipitations (in mm) recorded during a month is lower or equal to the double of the average temperature (in °C) of the same month. For the area considered, the more or less intense dry season lasts 10 months, from February to November over the period 1985–1999 but more than 11 months in the period 1999–2009 (Fig. 2). During this last period, the annual rainfall average (± standard deviation) was of 131.5 ± 74.5 mm. The variation coefficient thus exceeded 0.5, expressing the extreme uncertainty of precipitations, within the range of 47 mm observed in 2002 and 295 mm in 2004.

Ombrothermic diagram for Biskra.
2.2 Sampling
In order to estimate the quantitative composition of vegetation, three quadrats of 2 × 50 m separated by at least 10 m were randomly defined within each station. The number of individuals of each species was counted in each of these quadrats. The anthropised environments were studied in May 2010 and the steppes during the last week of April 2010. The plant species were identified using the following flora [22,23] and the nomenclature was updated from the tela-botanica website (http://www.tela-botanica.org). As for insects, each station was sampled once per month, or twice during the active period, from October 2008 until November 2009 for the oasian stations, and from May 2009 until April 2010 for the steppic stations. For each of the seven stations, the quantitative measure of acridian density was carried out using to a transect method. The total number of adult individuals belonging to each species was counted in a 100-m-long band of 1 m width. The countings in the bands were repeated three times for each station and day of sampling. The determinations were conducted using to the following references: [24–31] and the nomenclature updated from the OSF2 website (http://Orthoptera.SpeciesFile.org). Moreover, we consulted the collections of the MNHN of Paris thanks to the kindness of C. Amédégnato and S. Poulain, and the own personal of Dr. Jacques Mestre, in Sarlat. Three reference collections were assembled and kept at the Universities of Biskra, Constantine and Limoges.
2.3 Statistical processing
The variations of acridian assemblages according to the studied stations were analyzed by a Correspondences Analysis (CA), followed by an ascending hierarchical classification (AHC). Briefly, the scores obtained for the first three axes were used to calculate the Euclidean distances between taxa and stations. This method makes it possible to examine the differences in composition of samples [32]. We tested the total similarities between the sites by an ANOSIM (Analysis Of SIMilarity), a nonparametric test of significant difference based on a distance measure [33]. The index of Bray-Curtis was selected here as a measure of similarity and the P-values were deduced from 10 000 permutations. The contribution of each insect species to the differences observed between assemblages was calculated with the program SIMPER (SIMilarity PERcentage), using again the index of Bray-Curtis [33]. The significance of the differences in floristic diversity between two stations was calculated by a resampling procedure (bootstrap), according to the method developed by Poole [34]. All the multivariate analyses were conducted with PAST 1.98 [35].
3 Results
3.1 Floristic affinities between the seven stations
During the sampling of the seven stations, 61 species were recorded. For each station, we calculated the mean abundance of each species over the three quadrats carried out. The table obtained (Appendix I) was subjected to a CA presented in Fig. 3. One can recognize three quite separate units, each steppe medium being located on the right projection, while the five stations of oasis are concentrated in the left part, showing their strong homogeneity. In order to define the relative position of the stations taken in the oases, the five stations were submitted to another CA (Fig. 4) which reveals a projection fitting to a parabola, in the following order C1, A3, C2, A1 and A2, C1 being located towards the extreme left of the F1-F2 plane, while A2 being placed in the top right corner.

Ordination by CA of floristic samples in the seven studied stations.
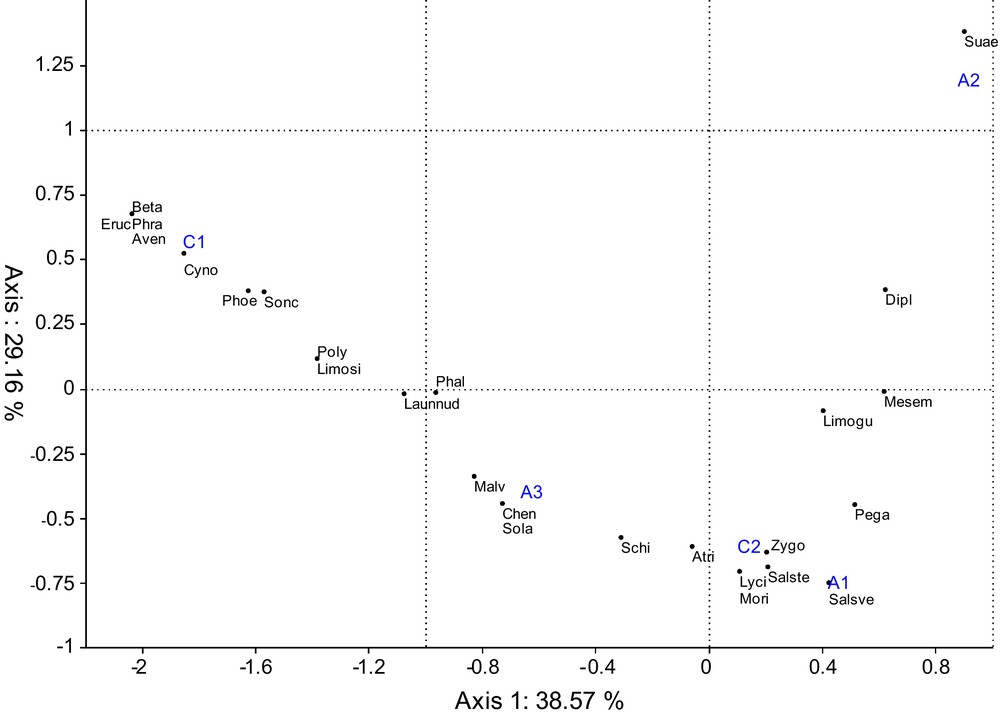
Ordination of the five stations in oases by CA (Abbreviation of species, see Appendix I).
3.2 Recognition of the acridian assemblages
Total numbers of 9291 acridian specimens, representing 45 species (Appendix II) were recorded in 288 quadrats, from which 96 synthetic samplings were established. This list increases considerably the knowledge previously acquired, since Tarai [6] noted 29 species and Harrat and Moussi [9] 20. It is highly probable that there were concerns in the determinations in the preceding inventories, mainly in the genera Dociostaurus (D. jagoi was reported whereas it is a new species for science [36]), and Sphingonotus. However, the collections assembled during 12 months in the seven stations made it possible to discover two new species for Algeria: Pamphagulus near to uvarovi, only known from Mauritania and Western Sahara [24], and Notopleura near to pygmaea, recorded in Tunisia [24]. The distribution of Aiolopus simulatrix, also known of Libya [37], is extended northward in Algeria as it was observed in Algerian Sahara, at Tamanrasset, Adrar and Ouargla [13], under the synonym A. savignyi (Krauss, 1890). Moreover, Hilethera aeolopoides, a Sahelian species poorly known in Algeria [38], is here well represented in the Biskra Wilaya, but rather rare at Bir-Naam. Among the 45 species, we removed five of the analyses because less than three individuals per species were observed: Schistocerca gregaria (Forsk, 1775), Aiolopus strepens (Latreille, 1804), Omocestus rufipes (Zetterstedt, 1821), Helioscirtus capsitanus (Saussure, 1884) and Locusta migratoria (L., 1758). The sampling-species (96 × 40) data set was subjected to a CA (Fig. 5) followed by an AHC based on the scores of the first three factors (Appendix III). According to the truncature line chosen, five units appear: the first includes the majority of the samplings taken within the sites A1, A2 and C2, the second gathers the samplings of A3 and C1 sites, the third and fourth relate to the samplings in the stony steppe, in spring on the one hand and in summer-autumn on the other hand. Finally, the fifth mostly consists of the samplings performed in sandy steppe. If the truncated line is moved towards the root, it also reveals two clusters of samplings within the sandy steppe, but the seasonal distribution is less obvious than for the stony steppe. To compare these clusters, we gathered all the samplings taken in the stony steppe, so as to obtain four assemblages.
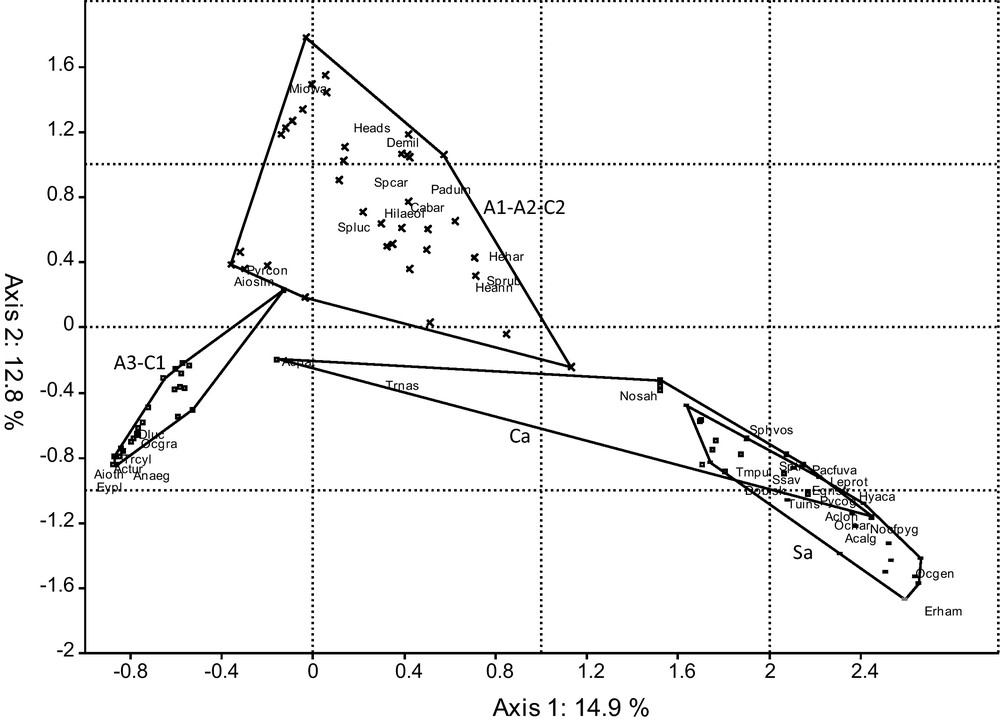
Ordination of Acridian assemblages in the oasian and steppic habitats. The code of species is given in Appendix II.
The difference in the composition of these four assemblages is supported by ANOSIM, since the Bonferroni corrected P-values, obtained after 10 000 permutations, are lower than 10−4, in all the combinations. Moreover, pairwise comparisons of Shannon H′ diversity index by bootstrap analyses emphasize significant differences (P < 10−3) between all the combinations, except in the case of A1A2C2 and Sa, where P = 0.102. As a result, the stony steppe has a diversity (H′ = 2.853) significantly higher than the sandy steppe (H′ = 2.541) and the A1A2C2 group (H′ = 2.351). The less diversified habitat is A3C1 (H′ = 1.902). On the other hand, the same analysis on these four assemblages with the richness S does not reveal significant differences, the values of S being between 20 and 24. The most characteristic species of each assemblage were deduced from SIMPER analysis (Table 2).
SIMPER analysis of the four assemblages. The characteristic species of each assemblage are in bold characters.
| Taxon | Contrib. | A1A2C2 | A3-C1 | Ca | Sa |
| Mioscirtus wagneri | 6.743 | 7.28 | 0.411 | 0 | 0 |
| Heteracris annulosa | 4.251 | 2.13 | 0.763 | 0.852 | 0.852 |
| Dericorys millierei | 3.581 | 3.09 | 0.0133 | 0.0183 | 0.0367 |
| Heteracris harterti | 2.678 | 1.81 | 0.0592 | 0.389 | 0.5 |
| Calliptamus barbarus | 2.582 | 2.6 | 0 | 0.314 | 0.0367 |
| Sphingonotus rubescens | 2.546 | 1.68 | 0 | 0.944 | 0.0183 |
| Heteracris adspersa | 2.397 | 3.34 | 0.0325 | 0 | 0.0183 |
| Hilethera aeolopoides | 2.204 | 1.92 | 0.371 | 0.222 | 0.0372 |
| Sphingonotus lucasi | 0.4856 | 0.521 | 0.17 | 0 | 0 |
| Pamphagulus bodenheimeri dumonti | 0.4536 | 0.2 | 0 | 0 | 0 |
| Sphingoderus carinatus | 0.03817 | 0.0308 | 0 | 0 | 0 |
| Ochrilidia gracilis | 11.78 | 0.252 | 20.8 | 0 | 0 |
| Duroniella lucasi | 8.067 | 0.0478 | 11.1 | 0 | 0 |
| Aiolopus thalassinus | 4.665 | 0 | 9.42 | 0 | 0.0183 |
| Acrotylus patruelis | 3.771 | 1.43 | 3.64 | 0.518 | 0.185 |
| Eyprepocnemis plorans | 1.071 | 0 | 2.71 | 0 | 0 |
| Acrida turrita | 0.6519 | 0 | 1.09 | 0 | 0 |
| Tropidopola cylindrica | 0.3538 | 0 | 0.873 | 0 | 0 |
| Anacridium aegyptium | 0.1236 | 0 | 0.352 | 0 | 0 |
| Pyrgomorpha conica | 10.76 | 6.81 | 8.88 | 0 | 0 |
| Aiolopus simulatrix | 0.03193 | 0.0267 | 0.0329 | 0 | 0 |
| Notopleura saharica | 3.049 | 0.872 | 0.0196 | 1.2 | 0 |
| Sphingonotus savigni | 2.004 | 0.035 | 0 | 1.87 | 0.0183 |
| Tuarega insignis | 0.7771 | 0 | 0 | 0.833 | 0.0372 |
| Sphingonotus vosseleri | 0.6763 | 0 | 0 | 0.593 | 0 |
| Tmethis pulchripennis | 0.6188 | 0.0567 | 0 | 0.407 | 0.0367 |
| Dociostaurus nov sp | 0.4937 | 0 | 0.00667 | 0.388 | 0 |
| Sphingonotus tricinctus | 0.3215 | 0 | 0 | 0.296 | 0 |
| Egnatioides striatus | 0.1083 | 0 | 0 | 0.111 | 0 |
| Eremogryllus hammadae | 2.728 | 0 | 0 | 0 | 1.72 |
| Ochrilidia geniculata | 2.349 | 0 | 0 | 0 | 1.24 |
| Ochrilidia harterti | 0.3714 | 0 | 0 | 0.0183 | 0.186 |
| Hyalorrhipis calcarata | 0.2886 | 0 | 0 | 0.0739 | 0.222 |
| Notopleura cf pygmaea | 0.2592 | 0 | 0 | 0.0183 | 0.148 |
| Acrotylus longipes | 3.203 | 0 | 0 | 1.68 | 1.09 |
| Acinipe algeriensis | 2.063 | 0 | 0 | 1 | 0.686 |
| Pyrgomorpha cognata | 2.052 | 0 | 0 | 0.944 | 0.612 |
| Pamphagulus cf uvarovi | 1.704 | 0 | 0 | 0.445 | 0.316 |
| Leptopternis rotschildi | 0.8834 | 0 | 0 | 0.278 | 0.444 |
| Truxalis nasuta | 0.5405 | 0.126 | 0.299 | 0.204 | 0.0183 |
3.3 Seasonal variations of the assemblages
The acridian assemblage of the stony steppes being split in two seasonal sub-assemblages, we wanted to know if this phenomenon is retrieved for the other studied habitats. ANOSIMs were thus carried out to compare the samplings performed during the spring period in a broad sense, i.e., from February until the beginning of June, relative to the rest of the year (Table 3). As it was expected, we found a significant difference for the stony steppes (P = 0.0084), but also for the sandy steppes (P < 10−4). If the two steppes are compared between them, season per season, there is a difference in composition in spring (P = 0.0006) but not in summer-autumn (P = 0.236), perhaps suggesting exchanges between the two habitats at this period. In the group A1A2C2 in oases, a seasonal difference is observed, but only marginally significant (P = 0.084). On the other hand, there exists a complete homogeneity in the group A3C1 (P = 1). If these two groups are compared between them, a significant difference in composition is observed in spring (P = 0.0018) and in summer-autumn (P < 10−4).
Comparison by ANOSIM between spring and summer-autumn faunas in the four habitats. The P-values were given by 10 000 permutations and Bonferroni-corrected.
| Ca Spring | Ca Summ-Aut | Sa Spring | Sa Summ-Aut | |
| Ca Spring | – | 0.0084 | 0.0006 | 0.0012 |
| Ca Summ Aut | 0.0084 | – | 0.0000 | 0.2358 |
| Sa Spring | 0.0006 | 0.0000 | – | 0.0006 |
| Sa Summ Aut | 0.0012 | 0.2358 | 0.0006 | – |
| A1A2C2 Spring | A1A2C2 Summ-Aut | A3C1 Spring | A3C1 Summ-Aut | |
| A1A2C2 Spring | – | 0.084 | 0.0018 | < 10−4 |
| A1A2C2 Summ Aut | 0.084 | – | 0.0006 | < 10−4 |
| A3C1 Spring | 0.0018 | 0.0006 | – | 1 |
| A3C1 Summ Aut | < 10−4 | < 10−4 | 1 | – |
The seasonal variations in the densities show larger amplitudes in oasian habitats than in steppic habitats (Fig. 6). The maximum densities are recorded in group A3C1 in July with approximately 180 individuals per 100 m2, i.e., about the double of the values measured in group A1A2C2. In the steppes, the values never exceed 30 individuals per 100 m2. With regard to the variations in richness (Fig. 7), the greatest species number is observed between October and May, with peaks more marked in the steppes than in the oases. In the two stations of steppe, we observe a fall of the richness at the beginning of May, more pronounced in the sandy steppe (from nine species in April, to three in May) than in the stony steppe (from 12 to 8). It is interesting to note that in the stony steppes, the richness in autumn is maintained better than in the other habitats.

Monthly variations of mean densities.
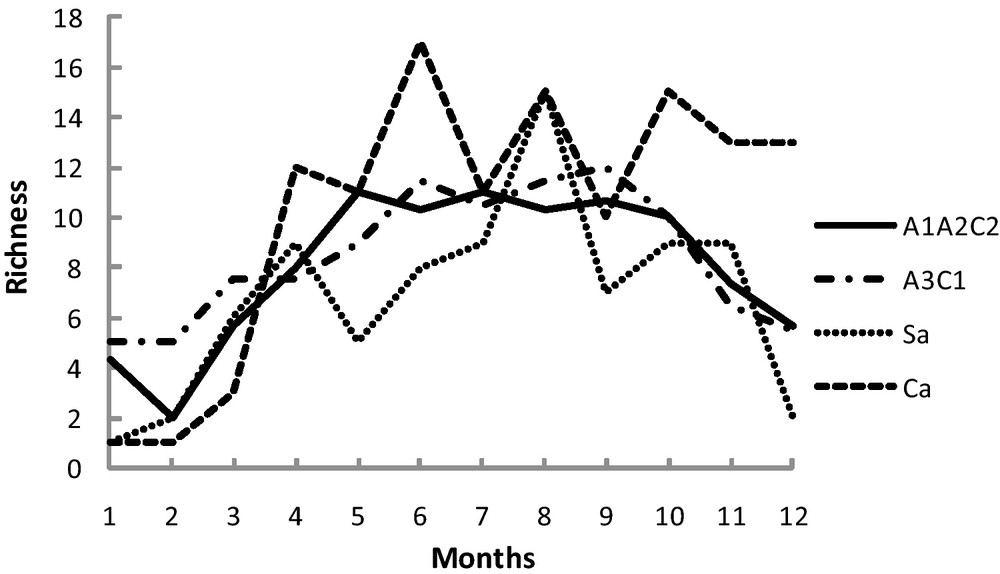
Monthly variations of richness.
4 Discussion
Biskra is a pre-Saharan area at the limit between the arid steppe and northern Sahara, as shown by the conjunction of several points: southern limit of the steppes with Alfa (S. tenacissima), northern limit of the perfect maturation of dates (P. dactylifera) and proximity to the isohyet 100 mm [22]. Now, the annual average of precipitations is about 131 mm, but it was around 144 mm in the period 1985–1999, 148 mm during the interval 1970–1985, and 156 mm between 1913 and 1938 (data from [39]). This area is thus still in the arid bioclimatic stage, but it can be extrapolated that it would belong to the Saharan bioclimatic stage within the next 30 years.
4.1 Interpretation of the vegetation (according to [20,40])
By the presence of S. tenacissima, L. spartum and P. albicans, the vegetation of both steppic stations belong to the phytosociological class of Lygeo-Stipetea Rivaz-Martinez 1978 em. Kaabeche 1990, i.e., more or less shrubby steppe, with an abundance of cespitous Poaceae (Sparte, Alfa) and annual species of irregular development. In spite of the predominance of Alfa in the sandy steppe and Sparte in the stony steppe, the floristic composition of both steppes belong to the same alliance, the Stipo-Launaeion Acanthocladae Kaabeche 1990, i.e., a steppe linked to the arid Mediterranean bioclimate, as attested by Helianthemum lippii, Thymelaea microphylla, Atractylis serratuloides, Cymbopogon schoenanthus and O. serrata. The vegetation developed in the sandy steppe is enriched with psammophytes species, such as Cleome amblyocarpa, Launaea mucronata, O. arenarium and Aristida obtusa. In spite of the presence of Sparte, this floristic association does not belong to the alliance including this species, because no other characteristic species are present. However, the presence of Pergularia tomentosa, Rhanterium adpressum, Farsetia aegyptia and Citrullus colocynthis indicates that this association has some characters close to the Pergularieto-Pulicarietea Quézel 1965, i.e., a vegetation growing on sandy, rocky or stony beds of the Sahara [41]. In summary, if the stony steppe is typically indicative of an arid Mediterranean bioclimate, the signification of sandy steppe is mixed in the sense that there are species characteristic of this same bioclimate and species assigned to the Saharan bioclimatic stage.
In the oasian stations, the group A1A2C2 has characters of halophilic frutescent and crassulescent vegetation, belonging to the class of Arthrocnemetea fruticosi Tüxen and Oberdorfer 1958, with the development of bushes of the Chenopodiaceae family, frequently found on the surroundings of salted depressions. As attested by S. vera, M. nodiflorum, A. halimus, S. tetragona, S. vermiculata, and Limoniastrum guyonianum, this group belongs to the order of Limonastretalia guyoniani Guinochet 1952. The group A3C1, as for it, is developed on salted ground (A3), even at little (C1), because of the reduction, or even of the absence of the majority of halophytes species. On the other hand, the ground is wetter there, especially in C1, as shown by the reed (Phragmites australis). The gradient of humidity expressed from A2, A1, C2, A3 towards C1 is explained by the absence of watering for the first two stations, of drip irrigation in the 3rd, of irrigation by seguias in 4th and a stronger irrigation in the last. Lastly, it is necessary to point out the presence of P. harmala and especially S. vermiculata in A1 station, which indicate an affinity to the class Pegano harmalae-Salsoletea vermiculatae Braun-Blanquet & Bolos 1957, developed on salted grounds with a nitrophilic and subnitrophilic character. One could interpret that by the course of ovine and caprine which enrich the ground with nitrogenized products.
4.2 Relationships between acridian fauna and vegetation
The most probable explanation about the acridian assemblages such as they are defined comes from the vegetation and not from a proximity effect. As shown in Table 1 and Fig. 1, the stations A1, A2 and A3 are geographically close to each other on the one hand, and ditto for C1 and C2 on the other. This does not fit to the A1-A2-C2 and A3-C1 acridian defined assemblages. Indeed, the five insect communities (four if the seasonal variations in the stony steppe are not taken into account) correspond to the floristic affinities of stations. The analysis of the vegetation in the oases makes it possible to better interpret the two acridian assemblages: the group A1-A2-C2 is associated with halophilic vegetation while the A3-C1 one is associated with irrigated cultures. It can thus be predicted that the first quoted assemblage should be also settled on the circumference of salted depressions, which represent abundant natural environments in steppic zones of Algeria. That could explain the relative richness of this assemblage. On the other hand, the A3-C1 assemblage is more pledged with the oasian cultures where the vegetation develops on a wetter ground. This more artificial character could give an account of the relative acridian poverty of this habitat. In summary, the oasian fauna does not result from a degradation of steppe fauna but constitutes a mixture of two original faunas.
The variations in the densities observed between the four assemblages are explained by humidity, since stations A3 and C1 exhibit the wettest vegetation and the highest acridian density, in total opposition with the habitats of steppe. Within A3-C1 assemblage, five species are clearly dominant, Ochrilidia gracilis, Duroniella lucasi, Aiolopus thalassinus, Acrotylus patruelis and Eyprepocnemis plorans. It should be noted that Pyrgomorpha conica is also abundant but this species is also found in the stations A1, A2 and C2. In this last group of stations, characterized by halophilic vegetation, the dominant acridians are Mioscirtus wagneri, Heteracris adspersa, Dericorys millierei and Calliptamus barbarus.
4.3 Phenology of the acridians according to the habitats
The observations carried out throughout the year on the adults do not make it possible to determine with certainty the type of cycle achieved by each species [42], but suggest results more or less supported by bibliographical data. The too rare species in a given habitat were not entered because they could result from migration from a close environment. That could concern Sphingonotus savignyi, Sphingonotus rubescens, H. aeolopoides, Truxalis nasuta, Tmethis pulchripennis, and Tuarega insignis, whose occurrence in sandy steppe is occasional and at a weak density compared to the stony steppe. In contrast, the rare presence of Ochrilidia harterti in stony steppe could be due to migrant individuals from the sandy steppe, but this would deserve to be tested by marking-recapture experiments, and the monitoring of instars. Bounechada et al. [5] gathered the species in the area of Sétif according to the stage which undergoes a winter diapause and also to the number of annual generations. We did not take again these categories but classified the species as a function of the number of generations and the period of maximum density, so as to test their adaptation toward the period of dramatic aridity. We considered three groups within the basic univoltine cycle, according to the period when the maximum of adults are observed. The first group, which is most abundant with 13 species, shows the highest density of adults in June and July (Table 4A). Then, if one places a limit in time around 25 June, the spring species present only one generation whose latest adults are recorded at this date. It contains Acinipe algeriensis, Eremogryllus hammadae (only on sand), Notopleura saharica (only on stones), and T. pulchripennis and T. insignis (especially on stones). In contrast, four species are observed as adult only in summer-autumn, and the first ones at this same date: Leptopternis rotschildi, Pamphagulus near to uvarovi, H. harterti, and O. harterti. Another phenology consists in the succession of two annual generations, the middle of the year seeing the end of the 1st generation and the beginning of 2nd. Owing to the two well-separated periods of adult occurrence, we arranged in this category the species: Dociostaurus novsp, Heteracris annulosa, D. millierei, Ochrilidia geniculata, Pamphagulus bodenheimeri dumonti, T. cylindrica, A. longipes, A. thalassinus, S. rubescens, T. nasuta and Notopleura near to pygmaea. Following [43], the cycle of Acrida turrita would comprise two to three generations in the littoral part of Algeria, which is compatible with our results. The phenology of A. patruelis in the area of Biskra seems similar to what is observed in the palm plantation of Ouargla, in the Algerian northern Sahara, with adults present almost all the year [13]. Consequently, the cycle would include at least three generations, in contrast to what is observed on the Mediterranean coastline where the larvae are recorded only in spring, indicative of a single generation [43]. The situation is comparable for the two species of Pyrgomorpha where the adults are observed all the year, as in the palm plantation in Ouargla [13]. In the area of Sétif, Bounechada and al. [5] argued that this species has only one annual generation with a larval diapause. Because of the annual distribution observed, we line up here with the conclusion of Ould El Hadj, but this problem deserves a closer examination.
Probable phenology of species. The different categories of cycles.
| Cycle 1-P | Acinipe algeriensis, Eremogryllus hammadae, Notopleura saharica, Tmethis pulchripennis, Tuarega insignis |
| Cycle 1-J | Calliptamus barbarus, Anacridium aegyptium, Eyprepocnemis plorans, Heteracris adspersa, Aiolopus simulatrix, Leptopternis rotschildi, Sphingonotus lucasi, S. tricinctus, S. vosseleri, Sphingoderus carinatus, Sphingonotus savignyi, Hyalorrhipis calcarata, Egnatus striatus |
| Cycle 1-S | Pamphagulus near to uvarovi, Ochrilidia harterti, Heteracris harterti |
| Cycle with 2 G | Dociostaurus novsp, H. annulosa, Dericorys millierei, Ochrilidia geniculata, O. gracilis, Pamphagulus bodenheimeri dumonti, Notopleura near to pygmaea, Tropidopola cylindrica, Acrotylus longipes, Truxalis nasuta, Aiolopus thalassinus, Sphingonotus rubescens |
| Cycle with 3 G | Acrotylus patruelis, Acrida turrita, Pyrgomorpha cognata, P. conica, Duroniella lucasi, Hilethera aeolopoides, Mioscirtus wagneri |
Probable phenology of species. Summary of phenology profiles according to the characteristic species of each assemblage (see Table 2).
| Species number | A1A2C2 | A3C1 | Sa | Ca |
| Cycle 1-P | 0 | 0 | 2 | 3 |
| Cycle 1-J | 5 | 3 | 2 | 5 |
| Cycle 1-S | 2 | 0 | 2 | 1 |
| Cycle with 2 G | 3 | 3 | 3 | 2 |
| Cycle with 3 G | 3 | 4 | 1 | 1 |
| Total | 13 | 10 | 10 | 12 |
| % 1-P/1-S/2 G | 38 | 30 | 70 | 50.0 |
| % 1-J | 38.5 | 30 | 20 | 41.7 |
In the natural environments, it is expected that the species present a phenology adapted to the aridity which prevails with an extreme intensity around the middle of the year, i.e. June and July. From the fundamental univoltine cycle centered on these two months, one can imagine several alternatives allowing attenuating the rigor of summer dryness: only one generation and shift towards spring or autumn, or two generations, one centered in spring and the other in summer-autumn. If these three categories are gathered (Table 4B), it appears that the sandy steppe is the habitat showing the highest corresponding proportion, reaching more than 2/3, while the lowest is observed in the irrigated part of oasis (A3-C1), which is less than a 1/3. Conversely, it is predicted that in the latter habitat, the cycle with at least 3 generations is most frequent, given the humidity linked to agriculture. Indeed, it is what we observe, as 40% of species share this phenology, in contrast to the sandy steppe habitat, which presents only 10% of this category. It thus seems that the sandy steppe is a habitat where the aridity is stronger than in the stony steppe, although both receive the same quantity of rain and at the same time. According to our analyses of the vegetation, it arises that the sandy steppe is richer in plant species with a Saharan affinity than the stony steppe. This stronger aridity of the sandy steppe could come from the sand, which would be drained more quickly than the ground of the stony steppe, and thus would offer worse conditions on arrival of the first heats. The replacements of sub-assemblages, more marked in the steppes than in the oasis, result in abrupt seasonal variations of the richness, as quoted previously. In particular, the reduction in the richness between April and May in the stony steppe is explained by the end of the 1st generation of several species, including Pyrgomorpha cognata. The decrease, even more marked in the sandy steppe, is due to the end of the 1st generation of O. geniculata and Notopleura near to pygmaea.
In conclusion, we highlighted within the oases two acridian assemblages, which are both different from those observed on the level of the steppes. In the wet biotopes of the oases, the most frequent phenology is the usual one retrieved in non-Mediterranean temperate areas. The assemblages of the steppes are characterized by a seasonal variation, in the sense that there exist two sub-assemblages, which we interpreted as the high frequency of phenology types adapted to mid-year aridity. If the climate shows tendencies for rainfall shortage, as in other parts of Algeria [44–46], one can expect a reduction of steppic landscapes and of the associated fauna characterized by its original life cycle and phenology.
Acknowledgements
We thank S. Poulain and the much regretted C. Amédégnato of the MNHN of Paris, J. Mestre de Sarlat, who enabled us to consult the collections of which they have the responsibility, A. Foucart and B. Massa for assistance in documentation. This work received financial support via the program Tassili 08MDU726 Globalbiodiv.