


 CC-BY 4.0
CC-BY 4.0
1. Introduction
Les méthodes d’imagerie ont révélé des architectures fonctionnelles d’aires cérébrales qui sont assez précises pour etre mathématisées.
Dans cet article1 , nous nous focaliserons sur des images mésoscopiques de V 1, la première des aires visuelles primaires. Bien sûr il existe de nombreux feedbacks descendants (« top-down ») des aires successives comme V 2 ou V 4 sur V 1. Mais V 1 est déjà très important. Selon la « high-resolution buffer hypothesis » de Mumford et Lee (Lee et al., 1998), V 1 n’est pas un simple « bottom-up early-module », mais participe à tous les processus visuels exigeant des résolutions fines. Son architecture fonctionnelle est par conséquent essentielle pour l’ensemble du système visuel.
Elle comprend trois grandes composantes, c’est-à-dire trois grandes classes de connexions. La première est celle des connexions « verticales » rétino-géniculo-corticales. La seconde et la troisième sont celles des connexions intracorticales : d’une part les connexions à courte portée et isotropes internes aux (hyper)colonnes d’orientation que nous allons définir, d’autre part les connexions « horizontales » cortico-corticales à longue portée et très fortement anisotropes entre (hyper)colonnes.
2. La structure hypercolumnaire de l’aire V1
Dans V 1, la densité de neurones est de l’ordre de 2 × 105/mm2 et les champs récepteurs sont de l’ordre de quelques degrés. Trois types de structures y interviennent, respectivement laminaire, rétinotopique et (hyper)columnaire. C’est la troisième qui nous intéresse ici. Elle est la grande découverte de Vernon Mountcastle et des prix Nobel (1981) David Hubel et Torsten Wiesel à la fin des années 1950.
Il existe dans V 1 des neurones « simples » (par opposition aux cellules « complexes » et « hypercomplexes ») sensibles à l’orientation, à la dominance oculaire et à la couleur. Leurs profils récepteurs sont connus (cf. par exemple DeAngelis, Ohzawa et al., 1995). Si l’on simplifie au maximum la situation en ne tenant compte ni de l’échelle (de la résolution et de la fréquence spatiale) ni de la phase, on peut dire que ces neurones détectent des couples (a,p) d’une position rétinienne a et d’une orientation p en a, ce que l’on appelle en géométrie des « éléments de contact ».
Par les méthodes d’enregistrement de réponses à des stimuli appropriés (barres orientées traversant le champ récepteurs des neurones, etc.), on a pu montrer que, perpendiculairement à la surface du cortex, la position rétinienne a et l’orientation préférentielle p restent à peu près constantes. Cette redondance « verticale » — qui définit un codage par population de la position — définit des colonnes d’orientation d’environ 20 μm. Comme l’a montré DeAngelis, Ghose et al. (1999) c’est la variation de la phase qui domine dans les colonnes. Par ailleurs, le codage par population permet au système d’avoir une résolution supérieure à celle des neurones individuels.
En revanche, le long de lignes parallèles à la surface du cortex, l’orientation préférentielle p varie par pas d’environ 10°. Un regroupement « horizontal » de colonnes d’orientation de même a variant de π définit une hypercolonne d’orientation qui est un micromodule neuronal large de 200 μm à 1 mm.
Le concept de colonne doit évidemment être notablement précisé et un nombre considérable de travaux lui ont été consacrés. Parmi les reviews disponibles sur ces sujets, on pourra consulter celle de Lund et al. (2003).
Nous nous occuperons d’espèces de mammifères, tupaya (tree shrew), chats, primates, hommes, où le concept de colonne d’orientation est d’une pertinence essentielle. Cela est assez restrictif. Toutefois, même en se restreignant à de telles espèces, il existe néanmoins une certaine diversité de la structure de V 1. La structure générale demeure invariante : rétinotopie, sélectivité à l’orientation, tuning de fréquences spatiales, etc., mais la structure fine est variable. Comme le note Van Hooser (2007),
« There is considerable diversity in the abundance of different cell classes, laminar organization, functional architecture, and functional connectivity. » 2
La structure hypercolumnaire de V 1 étant fondamentale, il est intéressant de trouver des mathématiques adéquates pour la modéliser. Un premier concept, tout à fait naturel, est, au niveau mésoscopique qui est le nôtre, le concept géométrique, bien connu des mathématiciens, de fibration.
3. V1 comme fibration mésoscopique
3.1. « Bridging scales » : le niveau mésoscopique
Nous nous situons à un niveau mésoscopique car les simulations « micro » réalistes d’une colonne d’orientation sont à elles seules d’une complexité inouïe et ont fait l’objet de certains parmi les plus gros projets computationnels mondiaux : le « Blue Brain Project » (lancé en 2005 par Henry Markram à l’École Polytechnique Fédérale de Lausanne) qui visait à simuler une colonne corticale de rat d’environ 10 000 neurones et 30 millions de synapses, puis en 2013, le « Human Brain Project » (flagship de l’Union Européenne à un milliard d’euros sur 10 ans, rival du projet américain « Brain ») qui en a pris le relai avec des supercalculateurs allant jusqu’au million de teraflops … .
3.2. Fibration et « engrafted variables »
Au niveau mésoscopique, à travers l’architecture fonctionnelle hypercolumnaire de V 1, à chaque position rétinienne a ∈ R du plan rétinien R se trouve associé de façon rétinotopique et anatomiquement observable un exemplaire (discrétisé) de l’espace P des orientations p du plan3 . P est codé par le cercle unité $\mathbb{S}^{1}$ des angles que font les orientations p par rapport à une orientation de référence et s’appelle traditionnellement en géométrie classique une « droite projective ». Il existe par conséquent une implémentation neuronale de la projection π : R × P→R du produit cartésien R × P sur son premier facteur R, projection qui est une « fibration » ayant pour base l’espace rétinien R et pour fibre la droite projective des orientations P.
La notion clé de fibration a été élaborée par les mathématiciens (puis, ensuite, par les physiciens théoriciens) pour modéliser des processus qui exigent d’associer à chaque point d’un espace de base M une entité d’un certain type F (un scalaire, un vecteur, un tenseur, une forme différentielle, une direction, une phase, un nombre quantique, etc.) dépendant « régulièrement » de ce point. Une solution évidente pour modéliser de tels champs sur M serait d’utiliser des applications 𝜑 : M→F. Mais dans bien des cas, il est nécessaire de considérer qu’à chaque point de M se trouve associé l’ensemble completF des valeurs possibles de telles 𝜑.
Très intuitivement, une fibration est constituée d’un espace de base M et de copies d’une variété F, appelée la fibre, « au-dessus » de chaque point de F (cf. Figure 1). Globalement, l’espace E de la fibration (les fibres recollées ensemble) n’est pas forcément le produit cartésien M × F. Il résulte du recollement de plusieurs produits cartésiens Ui × F définis sur des domaines locaux Ui de M (cf. Figure 2).
Le schéma général d’une fibration d’espace de base M, de fibre F et d’espace total E. Au-dessus de chaque point x de M la fibre π−1(x) = Ex est isomorphe à F.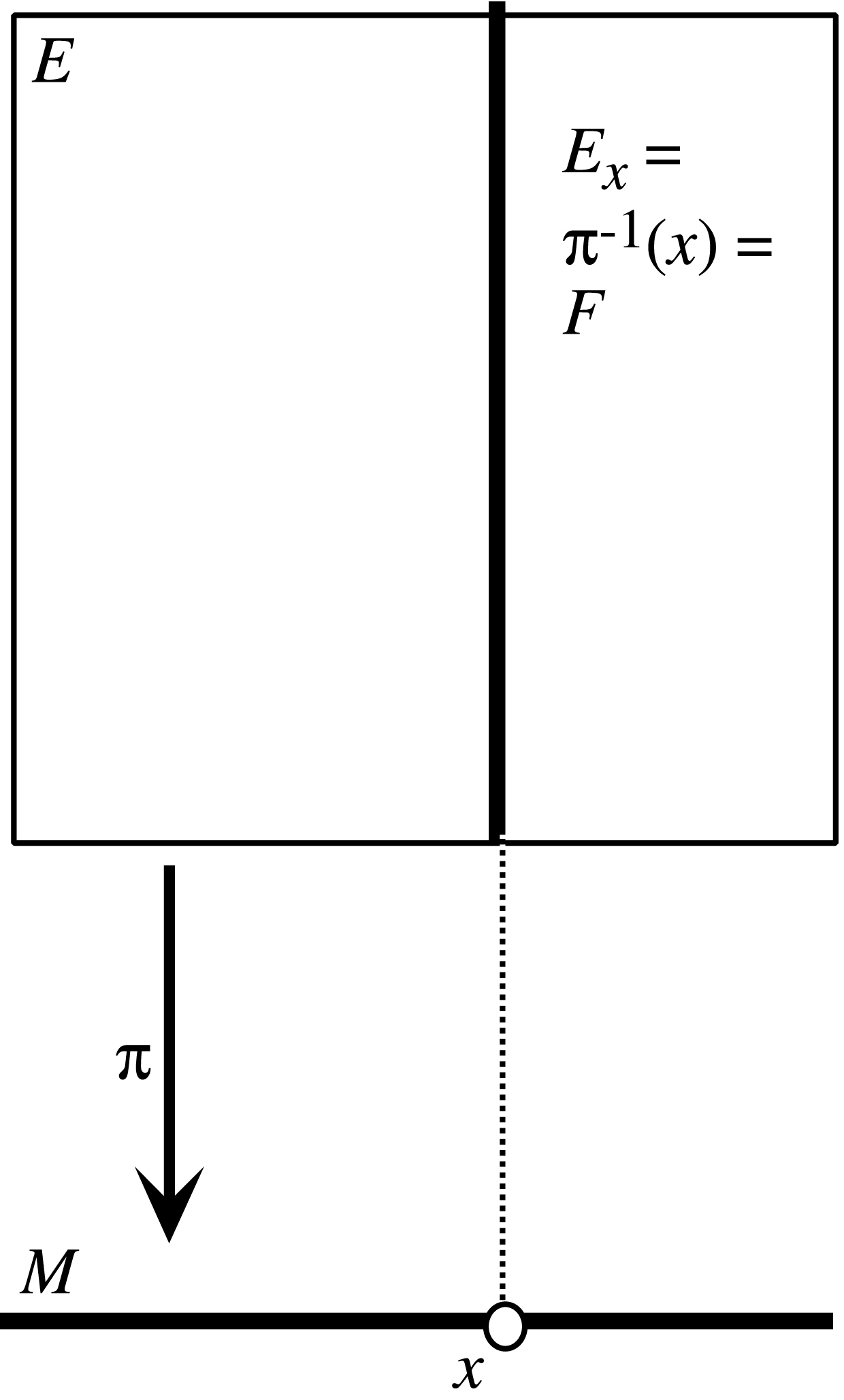
La trivialité locale d’une fibration. Pour tout point x de M il existe un voisinage U de x dont l’image inverse π−1(U) = EU est le produit direct U × F, π étant la projection sur le premier facteur.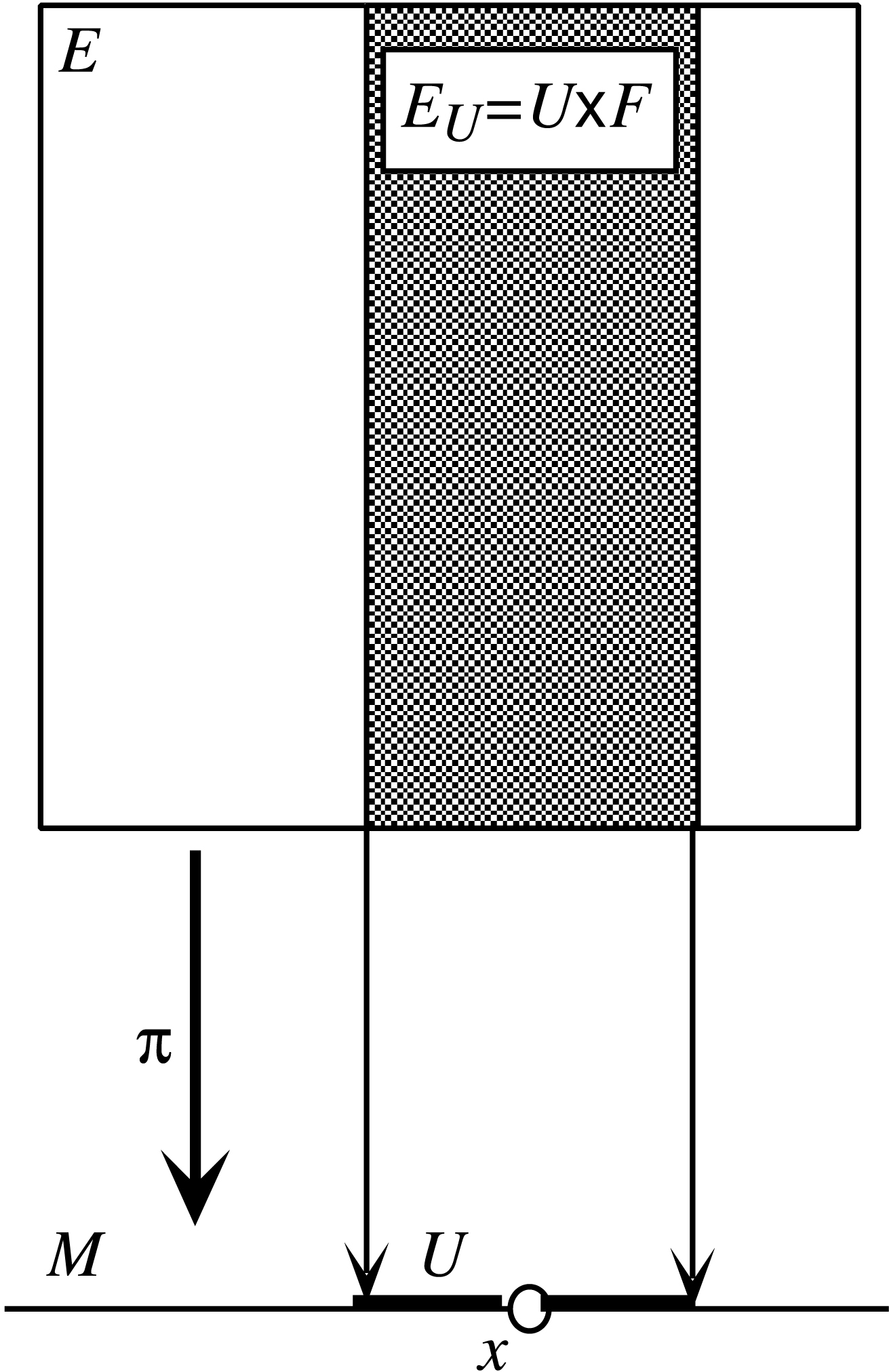
Une section d’une fibration est une application différentiable qui relève la projection π, en associant à chaque point x de la base M un élément de la fibre Ex. Si s : M→E est une section, l’application composée π°s est donc égale à l’identité de M (cf. Figure 3).
Une section d’une fibration définie sur un ouvert U de M associe à chaque point x de U une valeur s(x) dans la fibre Ex au-dessus de x.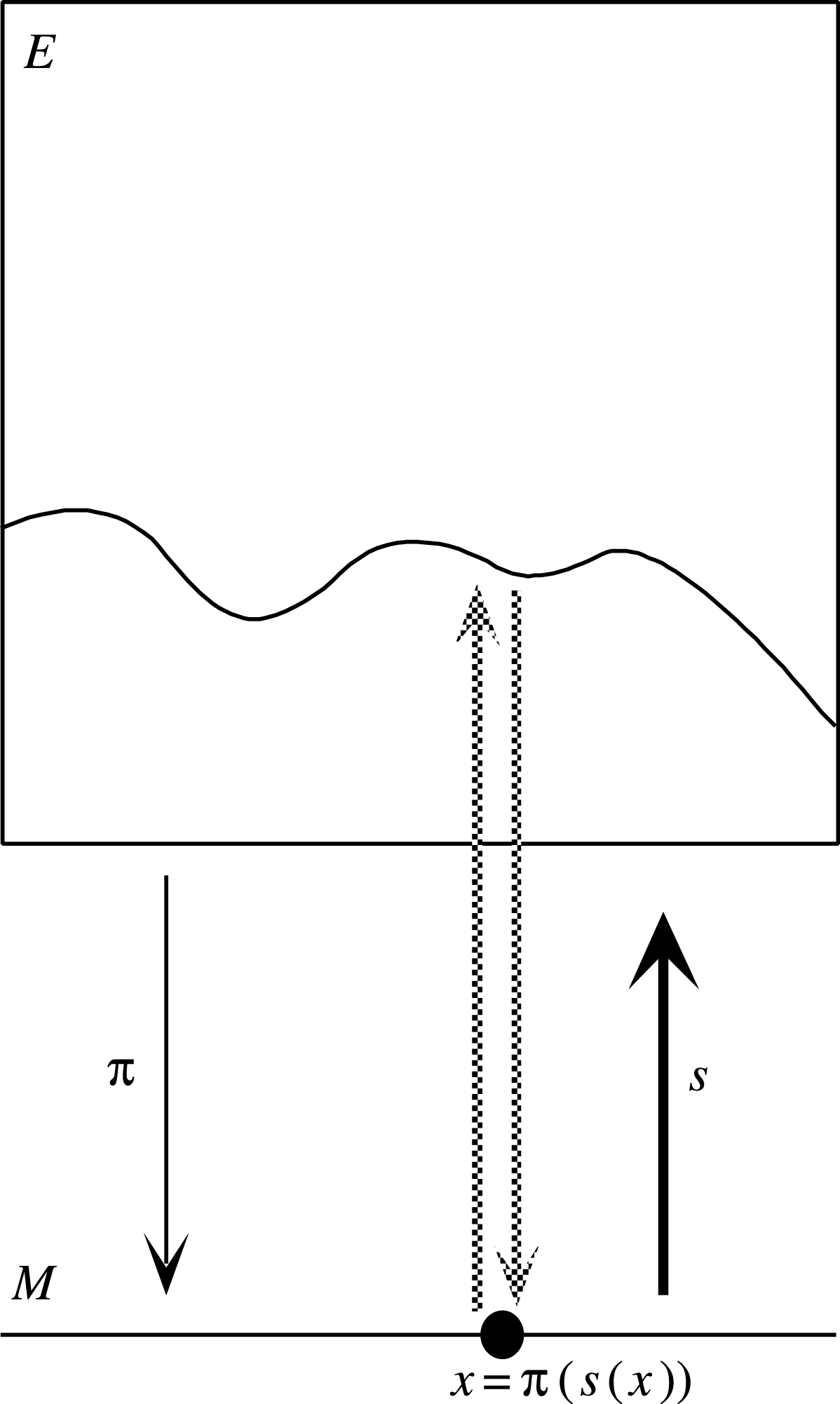
Dans notre cas la projection π : R × P→R est mathématiquement triviale mais ne l’est pas du tout neurophysiologiquement :
- les champs récepteurs des neurones des colonnes corticales sont des très petites cartes locales s’intersectant et se recollant entre elles ;
- il n’est pas du tout évident de tester expérimentalement la structure de produit direct, c’est-à-dire l’indépendance des variables de position et d’orientation ; les différences interspécifiques sont importantes : pour certaines espèces c’est une même couche de V 1 qui implémente R × P alors que pour d’autres ce sont des couches différentes ;
- la projection (au sens géométrique) π est implémentée par toute la fine connectique des voies rétino-géniculo-corticales qui projettent (au sens neurophysiologique) la rétine sur V 1 4 .
Quoi qu’il en soit de ces difficultés, dans les modèles mésoscopiques présentés ici, une colonne avec ses connexions internes isotropes à courte portée se réduit à un simple élément de contact (a,p) et une hypercolonne, avec également ses connexions internes, se réduit à une fibre de π de forme {a} × P. Cela peut sembler être une réduction inacceptable eu égard aux projets computationnels que nous venons d’évoquer. Mais notre propos est de montrer que, malgré cette énorme réduction de leur complexité, l’organisation fonctionnelle de ces éléments conduit cependant déjà à une notable complexité.
La structure géométrique constituée par le produit de R comme espace de base avec un espace de variables « secondaires » (orientation, dominance oculaire, couleur, direction du mouvement, etc.) a été fort bien formulée par David Hubel avec le concept d’« engrafted variables » :
« What the cortex does is map not just two but many variables on its two-dimensional surface. It does so by selecting as the basic parameters the two variables that specify the visual field coordinates (distance out and up or down from the fovea), and on this map it engrafts other variables, such as orientation and eye preference, by finer subdivisions. » Hubel (1988, p. 131)
C’est ce modèle de V 1 proposé par Hubel qui nous a conduit au concept géométrique fondamental de fibration ou d’espace fibré.
Toutefois il faut tenir compte d’une contrainte dimensionnelle. La fibration π : R × P→R est abstraitement de dimension 3 (2 degrés de liberté pour la position rétinienne a = (x,y), 1 degré de liberté pour l’orientation p), alors que les couches corticales sont essentiellement de dimension 2. Il existe par conséquent un problème de « collapse dimensionnel ». Les systèmes visuels issus de l’évolution qui nous intéressent ici l’ont résolu à travers la fascinante structure en « pinwheels » (roues d’orientation) de V 1 analysée en détail depuis les travaux pionniers de Tobias Bonhöffer, Gary Blasdel et Amiram Grinvald.
4. La structure en pinwheels de V1
4.1. L’observation des pinwheels
4.1.1. Les cartes fonctionnelles d’orientation
Les expériences permises par les progrès de l’imagerie cérébrale ont montré que les hypercolonnes sont en fait géométriquement organisées en « roues d’orientation » baptisées « pinwheels ». La couche corticale observée est réticulée par un réseau de points singuliers (distants d’environ 1200 μm chez le chat et d’environ 600 μm chez le primate) qui sont les centres de pinwheels locaux qui se recollent en une structure globale. La méthode d’imagerie employée a été mise au point au début des années 1990, entre autres par Bonhöffer and Grinvald (1991). Baptisée « in vivo optical imaging based on activity-dependent intrinsic signals », elle est fondée sur le fait que l’activité métabolique des tissus nerveux change leurs propriétés optiques et elle permet d’acquérir ainsi des images de l’activité des couches corticales superficielles. Plus précisément, elle repose sur l’absorption différentielle d’oxyhémoglobine ou de déoxyhémoglobine, ou sur des colorants (dyes) dont la fluorescence est un indicateur de la dépolarisation locale des neurones.
Cette méthode a amorcé une véritable révolution. Elle a permis de visualiser les organisations fonctionnelles. Comme le soulignent Ohki and Reid (2007),
« optical imaging revolutionized the study of functional architecture by showing the overall geometry of functional maps. »
Mais le défi expérimental est considérable. D’abord le rapport signal/bruit est très petit (∼10−3) car le bruit de fond est énorme. Ensuite des millions de neurones sont connectés entre eux, chacun possédant des centaines (voire des milliers) de synapses et l’imagerie opère à une méso-échelle définie par des moyennages (mais il existe des méthodes de microscopie confocale biphotonique de niveau « micro »). Un « neurone » méso est en fait une position corticale où se trouve localisé un paquet de neurones. Qui plus est, les méthodes d’imagerie optique in vivo qui ont une bonne résolution spatiale mésoscopique (50 μm) ont en revanche une mauvaise résolution temporelle. Pour visualiser la dynamique corticale on a besoin d’autres méthodes comme les colorants voltage-dépendants (voltage-sensitive dyes) qui permettent d’obtenir des résolutions temporelles de l’ordre de la ms.
Pour une présentation de ces nouvelles techniques d’analyse du cerveau des mammifères, on pourra se référer à la synthèse (Grinvald et al., 2001) présentée par son créateur Amiram Grinvald.
Dès la fin des années 80, de telles cartes ont commencé à être disponibles. Dans un protocole standard, on présente à l’animal plusieurs dizaines de fois (de 20 à 80 fois) des grilles (gratings) de fort contraste constituées de bandes noires (par exemple de 6.25°) alternées avec des bandes blanches (par exemple de 1.25°), avec plusieurs (par exemple 8) orientations différentes et une vitesse par exemple de 22.5°/s. On ouvre une fenêtre crânienne au-dessus de V 1 et on illumine le cortex en lumière orange (605 nm). On observe alors, en fonction de l’orientation des gratings, des patterns d’absorption différentielle dus aux inhomogénéités spatiales locales du rapport déoxyhémoglobine / oxyhémoglobine. On somme ensuite les images de l’activité de V 1 obtenues pour différentes grilles et l’on construit des cartes différentielles que l’on normalise et dont on élimine le bruit de basse fréquence.
On obtient ainsi des cartes fonctionnelles comme celles de la Figure 4 et de la Figure 5 dues à Crair et al. (1997) et Bosking et al. (1997) et concernant les couches 2/3 d’un tree shrew (tupaya) : LGN → couche 4 → (strictement feedforward) → couches 2/3. (Le tupaya ressemble aux primates mais, contrairement à eux, n’a pas de distinction fovéa/périphérie, ce qui facilite les observations). Les orientations y sont codées par des couleurs et les lignes d’iso-orientation y sont donc les lignes monochromatiques.
Méthode d’établissement d’une carte d’orientation de V 1 (les orientations préférentielles sont conventionnellement codées par des couleurs). (D’après Crair et al., 1997).
V 1 d’un tree shrew (tupaya). Les différentes orientations sont codées par des couleurs. À droite, zoom sur des exemples de points réguliers et de points singuliers de chiralités opposées. (D’après Bosking et al., 1997).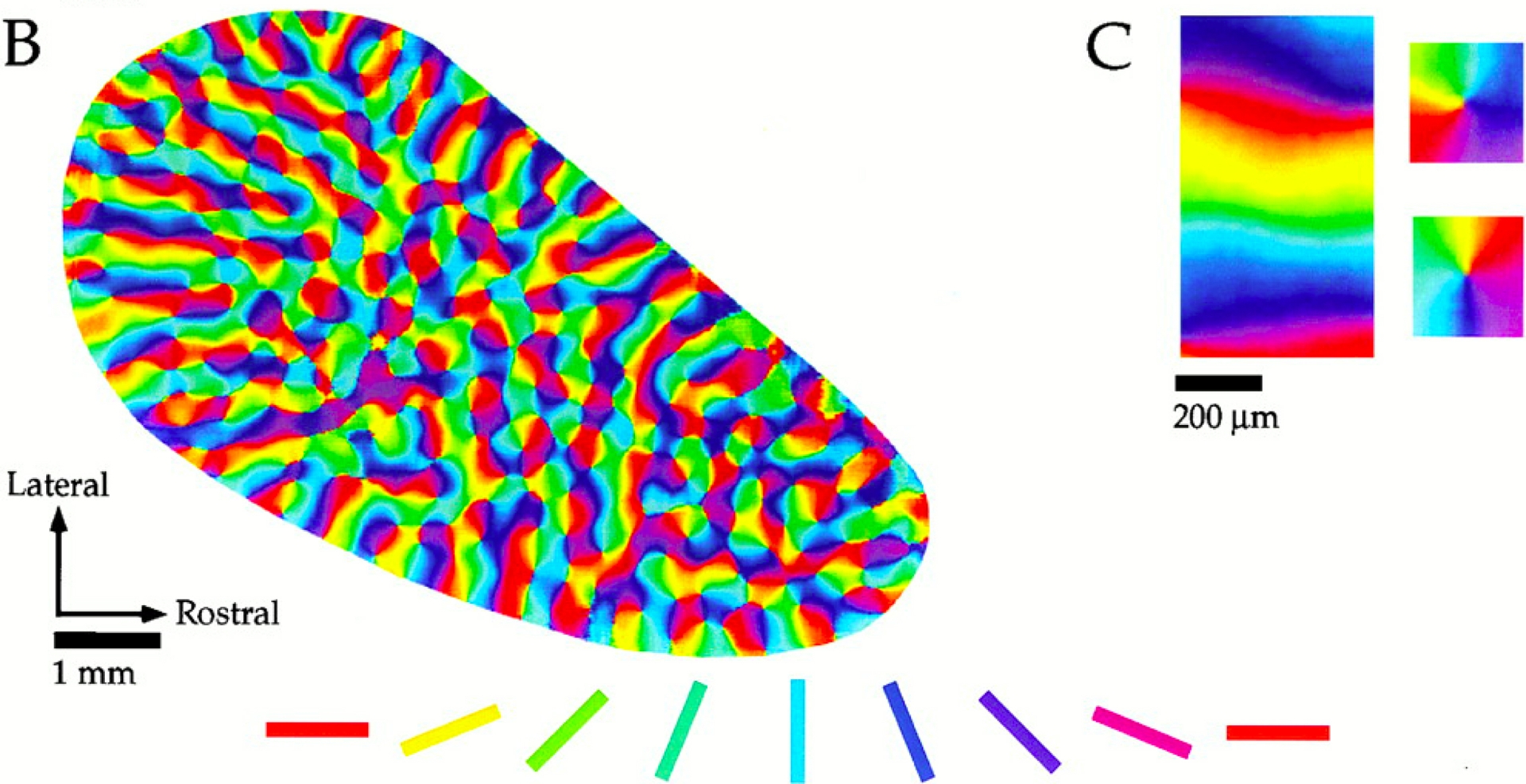
On remarque qu’il existe trois classes de points :
- Des points réguliers où le champ d’orientation est localement trivial au sens où les lignes d’iso-orientation y sont approximativement parallèles.
- Des points singuliers au centre des pinwheels où convergent toutes les orientations. Ils ont une « chiralité » positive ou négative et sont de chiralités opposées lorsqu’ils sont adjacents.
- Des points cols au centre des cellules du réseau, points où les lignes d’iso-orientation bifurquent.
La Figure 6, basée sur la figure 14 du chapitre 27 “Central Visual Pathways” du traité classique Kaschube et al. (2008), montre mieux les lignes isochromatiques qui sont les lignes de champ d’iso-orientation de V 1.
Les lignes isochromatiques sont les lignes d’iso-orientation de V 1. (D’après Kaschube et al., 2008).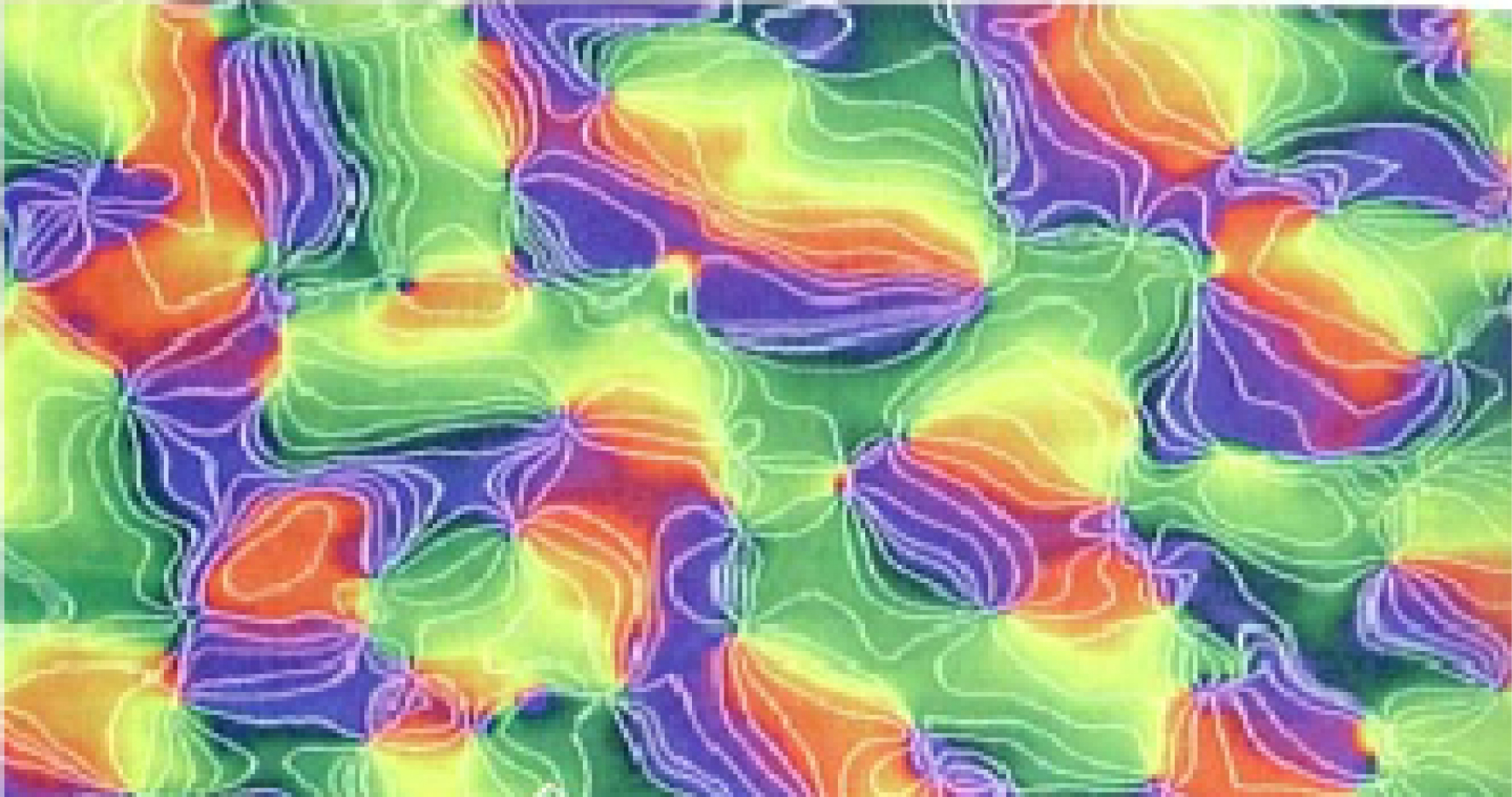
Depuis ces travaux pionniers, de nombreux approfondissements ont été obtenus, en particulier avec des stimuli qui sont des images naturelles (cf. par exemple l’article de 2004 de Wang et al., 2024). Mais dans cet article nous nous limiterons aux cartes fonctionnelles de base.Remarque.
4.1.2. 2D − 2(1/2)D − 3D
Les cartes d’orientations avec leurs « engrafted variables », leurs pinwheels et leurs singularités ci, sont des entités géométriques fort intéressantes ayant le statut de « fibrations partielles ». Au-dessus des points singuliers ci on a la fibre des orientations $P=\mathbb{S}^{1}$. Mais au-dessus des points réguliers on a seulement une orientation. Une carte d’orientation sans pinwheels serait un simple champ d’orientation régulier et serait une entité de dimension 2. Une fibration complète avec un exemplaire de P au-dessus de chaque point serait une entité de dimension 3. Les cartes d’orientation sont donc si l’on peut dire des entités de dimension « 2(1/2) ».
Il y a donc une balance, mesurée par la densité des pinwheels, dans la façon dont les cartes représentent concrètement (neurophysiologiquement) la variété abstraite 3D des éléments de contact (a,p) sur une couche 2D. Une carte est une approximation discrétisée 2(1/2)D de la fibration 3D. D’où l’intérêt d’étudier plus précisément la structure fine des cartes au voisinage de leurs singularités.
4.1.3. La structure des cartes au voisinage des singularités
En couplant d’une part des méthodes d’imagerie et d’autre part des méthodes d’enregistrement intracellulaire de spikes déclenchés par les inputs synaptiques et de potentiels de membrane de neurones individuels, Maldonado et al. (1997) ont analysé la structure fine des cartes d’orientation aux singularités. Ils ont constaté que
“orientation columns contain sharply tuned neurons of different orientation preference lying in close proximity.” (p. 969)
Autrement dit, la redondance colomnaire disparaîtrait aux points singuliers.
Par ailleurs, Schummers et al. (2002), en utilisant comme stimuli des grilles en mouvement, ont montré de même que
“neurons near pinwheel centers have subthreshold responses to all stimulus orientations but spike responses to only a narrow range of orientations.” (ibid., p. 969)
On évalue la sélectivité des réponses spikantes et des réponses sous-liminaires en fonction de la position par rapport aux pinwheels. Loin des pinwheels, les cellules
“show a strong membrane depolarization response only for a limited range of stimulus orientation, and this selectivity is reflected in their spike responses”. (ibid., p. 970)
Au centre d’un pinwheel, au contraire, seule la réponse spikante est sélective et il existe une forte dépolarisation de la membrane pour toutes les orientations.
Cela laisse supposer qu’aux points singuliers toutes les orientations sont bien en quelque sorte présentes. C’est une solution originale au problème des singularités qui renvoie à la connectivité des micro-circuits neuronaux mis en jeu.
“These examples indicate that both simple and complex cells located near pinwheel centers receive synaptic inputs over a broad range of stimulus orientations, although not all of these inputs are represented in the spike outputs.” (ibid., p. 971)
En résumé,
“Neurons close to pinwheel centers receive inputs at all stimulus orientations, whereas neurons far from pinwheel centers only receive synaptic inputs over a narrow range of orientations.” (ibid., p. 974)
4.1.4. Microscopie confocale biphotonique
Mais les techniques d’imagerie évoquées jusqu’ici ne sont pas assez précises. Aujourd’hui de nouvelles techniques comme celles de l’imagerie in vivo par microscopie confocale biphotonique ont fourni des cartes fonctionnelles ayant une résolution au niveau des neurones individuels. Kenichi Ohki et ses collègues (Ohki, Chung et al., 2006) ont montré que, chez le chat, les pinwheels définis au niveau méso restent très bien ordonnés au niveau micro et que par conséquent
“pinwheels centers truly represent singularities in the cortical map”.
La méthode consiste en l’injection d’indicateurs calciques (Oregon Green BAPTA-1 acetoxylmethyl esther) qui étiquettent quelques milliers de neurones dans des régions de taille 300–600𝜇. On mesure simultanément les signaux calciques évoqués par les stimuli visuels dans des centaines de neurones à différentes profondeurs (de 130 à 290𝜇 par pas de 20𝜇). Comme le montre la Figure 7, on trouve des pinwheels avec la même roue d’orientation.
“This demonstrates the columnar structure of the orientation map at a very fine spatial scale.”
L’imagerie optique biphotonique mesure simultanément les signaux calciques évoqués par les stimuli visuels dans des centaines de neurones à différentes profondeurs (de 130 à 290𝜇 par pas de 20𝜇). On trouve des pinwheels analogues possédant la même roue d’orientation à ces différentes profondeurs. Ohki, Chung et al. (2006).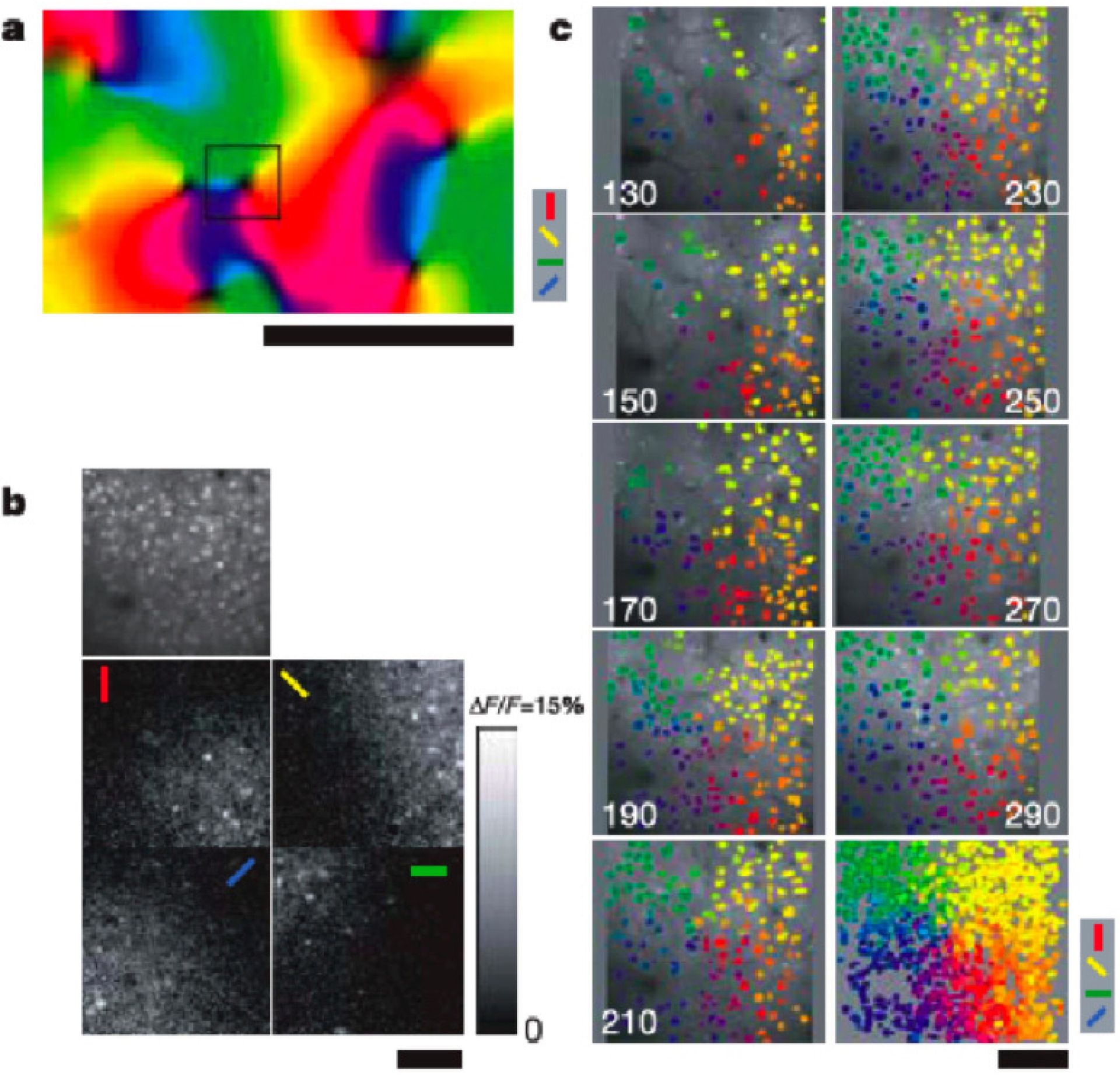
D’où le problème de la connectivité implémentant la sélection fine des orientations près des singularités. Plusieurs hypothèses sont envisagées par les auteurs concernant l’arbre dendritique près d’un centre C (quelques dizaines de 𝜇) dans un domaine D d’iso-orientation :
- arbre dendritique déséquilibré vers D ;
- arbre dendritique symétrique, mais inputs excitateurs déséquilibrés vers D ;
- arbre dendritique symétrique, inputs excitateurs symétriques mais locaux et intérieurs à D (bonne ségrégation au voisinage de C) ;
- arbre dendritique symétrique, inputs excitateurs symétriques et intégrés sur une grande aire dendritique.
4.1.5. Périodicité
Dans la Figure 5 on remarque qu’il existe une sorte de longueur caractéristique — une maille, un nombre d’onde — du réseau. On peut mesurer de façon précise cette périodicité en prenant la carte des pinwheels, en la translatant de t = (u,v) et en calculant la corrélation des deux cartes. L’autocorrélation C(t) a évidemment un pic primaire en t = (0,0) (la carte s’auto-corrèle parfaitement avec elle-même) et des pics secondaires donnent la périodicité.
En cas d’isotropie, C(t) ne dépend que du module r = |t| de t. Si l’on considère alors la transformée de Fourier P(k) de C(r) (ce que l’on appelle le « spectre de puissance »), on constate que P(k) est maximal sur un anneau de rayon k0 = 2π/Λ0 et Λ0 définit la maille du réseau de pinwheels.
La Figure 8 due à Niebur and Wörgötter (1994) montre une carte d’orientation chez le macaque (aire 18) et le spectre de puissance concentré sur un anneau de rayon moyen k0 = 2π/Λ0. Nous allons revenir plus bas sur le cas limite où le spectre de puissance est totalement concentré sur le cercle de rayon k0.
La périodicité d’une carte d’orientation (à droite). Le spectre de puissance (la transformée de Fourier de la fonction d’autocorrélation de la carte) est concentré sur un anneau de rayon moyen k0 = 2π/Λ0 et Λ0 définit la périodicité de la carte. (D’après Niebur and Wörgötter, 1994).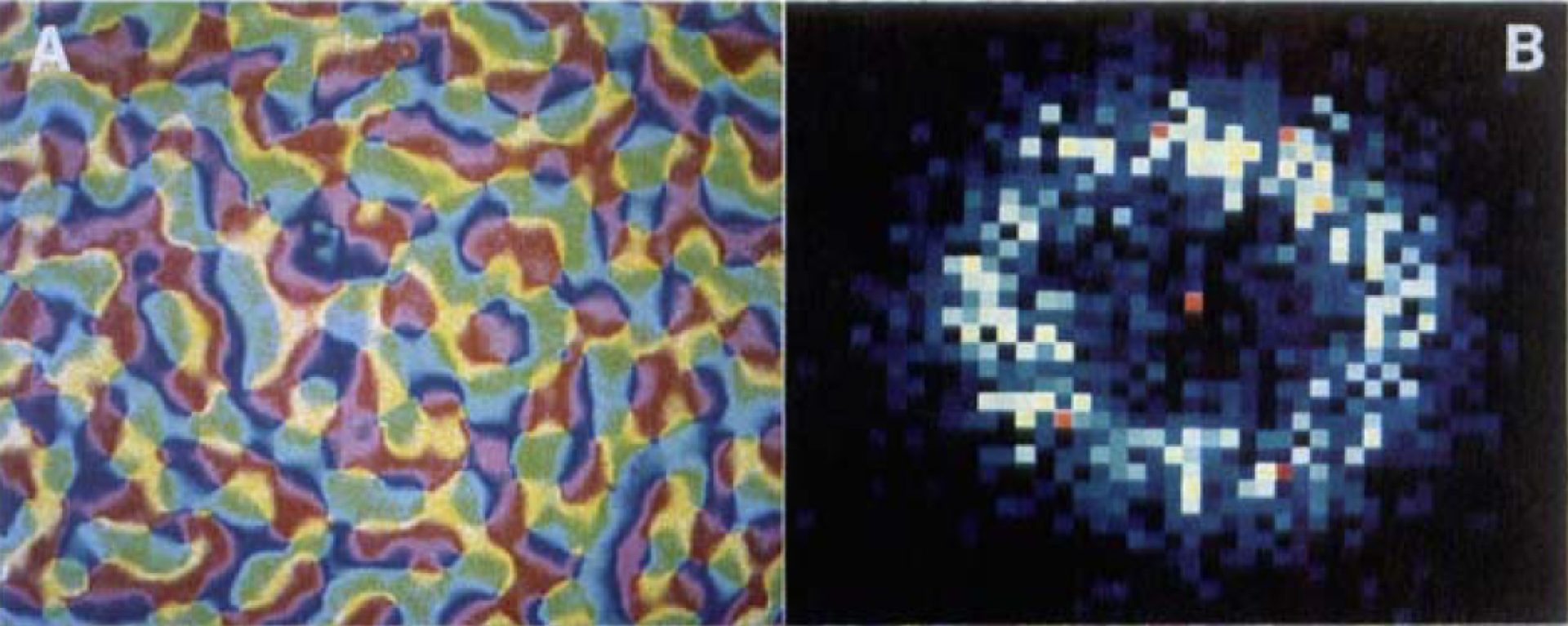
Cette périodicité est utilisée dans des modèles « cristallographiques » des cartes où les pinwheels sont distribués sur des réseaux réguliers (carrés, hexagonaux, etc.) possédant de fortes symétries (cf. par exemple notre article de 2003 (Petitot, 2003)). Une hypothèse de symétrie permet de mieux calculer la dynamique des activités de V 1. Un bon exemple est l’article (Veltz et al., 2015) de Romain Veltz, Pascal Chossat et Olivier Faugeras.
La structure la plus normale de V 1 est ainsi une distorsion de réseaux relativement « cristallins », ce qui correspond au fait que les chiralités de pinwheels adjacents sont opposées. Mais la distorsion peut devenir considérable au bord entre V 1 et V 2 car, comme l’ont montré entre autres Kenichi Ohki et des collaborateurs dans Ohki, Matsuda et al. (2000), des pinwheels de même chiralité sont alignés le long du bord (cf. Figure 9).
La disposition des pinwheels au voisinage de la frontière entre V 1 et V 2. Elle est très distordue car elle comprend des alignements de plusieurs pinwheels de même chiralité. (D’après Ohki, Matsuda et al., 2000).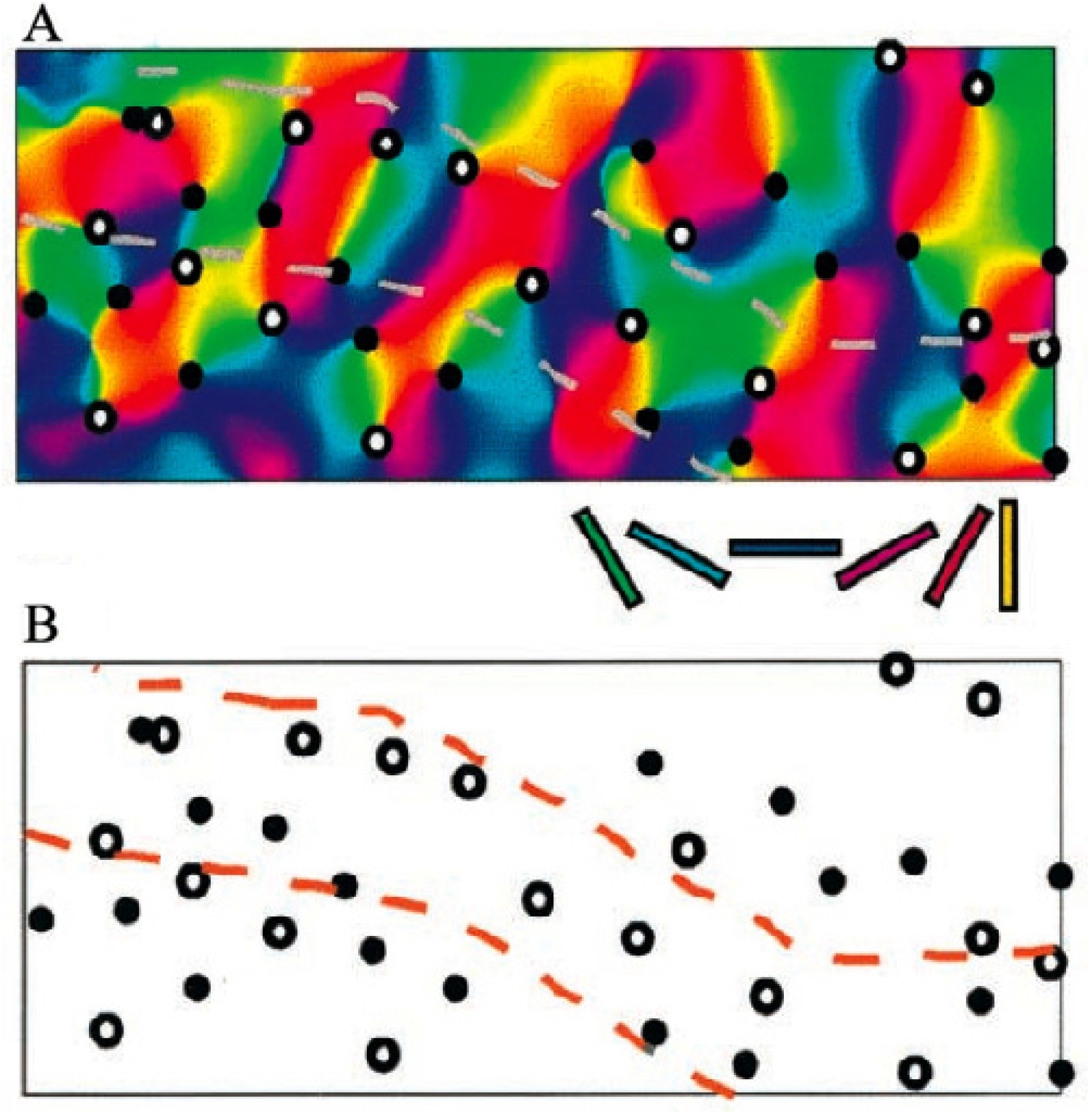
4.2. Les cartes fonctionnelles comme champs: le « spatial layout »
Il est intéressant de noter que les réseaux de pinwheels avec leurs lignes d’iso-orientation ressemblent à des modèles de type “champ”. Nous entendons ici par “champ” la donnée, en (presque) chaque point d’un espace, d’une direction ou d’un vecteur tangent variant régulièrement avec le point. Les lignes de champ sont alors les courbes tangentes en chacun de leur point à la direction ou au vecteur tangent donné en ce point. Quant aux singularités ce sont les points exceptionnels où le champ s’annule ou bien où plusieurs directions sont présentes.
4.2.1. L’abduction de Braitenberg
Ici, les singularités ci situées aux centres des pinwheels sont analogues, avec leur chiralités, à des charges ± produisant des lignes de champ dans $\mathbb{R}^{2}$. Une telle structure de champ a été introduite très tôt par Valentino et Carla Braitenberg, en fait dès 1979, bien avant l’introduction des méthodes d’imagerie optique in vivo, dans V. Braitenberg and C. Braitenberg (1979). Partant des résultats de 1962 de Hubel et Wiesel sur le singe et le chat, ils ont anticipé par de remarquables inférences abductives l’organisation radiale en pinwheels découverte plus tard expérimentalement.
4.2.2. L’abduction de Swindale
Dans un papier étonnant de 1987 (Swindale et al., 1987) (toujours avant l’apparition des techniques d’imagerie optique), Nicholas Swindale, Joanne Matsubara et Max Cynader ont reconstruit abductivement, pour l’aire 18 du chat, la première carte d’orientation 2D (bidimensionnelle) présentant des pinwheels. Il s’agit d’une vraie performance. Ils surent interpoler entre les orientations préférentielles mesurées à différents sites et construire une carte « fine grained » représentée à la Figure 10.
La reconstruction par Swindale d’une carte d’orientation 2D « fine grained » à partir d’enregistrements d’électrodes. (D’après Swindale et al., 1987).
Ils découvrirent alors les fameux futurs pinwheels qui confirmaient l’organisation radiale anticipée par les Braitenberg et qu’ils appelèrent des « 180° singularities » parce que l’orientation tourne de π lorsque l’on tourne de 2π autour du point singulier. En introduisant un code-couleur pour les orientations, ils obtinrent alors une carte d’orientation analogue à celles construites plus tard par imagerie in vivo.
Mais les auteurs allèrent d’emblée plus loin. Ils comprirent la structure du champ des orientations près des singularités. Ce sont soit des points d’arrêt soit des points triples.
4.3. Points d’arrêt et points triples
Il est essentiel de bien comprendre qu’il existe deux champs dans une carte d’orientation, dédoublement qui donne à ces cartes leur intérêt géométrique.
- le champ dont les lignes de champ sont les lignes isochromatiques, i.e. les lignes d’iso-orientation ;
- le champ dont les lignes de champ sont les intégrales du champ d’orientations lui-même.
Ces deux champs sont différents car en général l’orientation p en un point a n’est pas du tout tangente à la igne d’iso-orientation passant par a. Qui plus est, le premier champ est défini modulo 2π et le second modulo π.
La Figure 11 est due à Shmuel and Grinvald (2000). Nous y avons ajouté (en noir) les lignes du champ d’orientation au voisinage de deux singularités de chiralités opposées.
Cartes des orientations et des pinwheels du V 1 d’un tupaya. On y observe la relation entre les pinwheels (couleurs) et les orientations préférentielles. On a représenté en noir les lignes de champ d’orientation au voisinage de deux singularités de chiralité opposée. (En partie d’après Shmuel and Grinvald, 2000).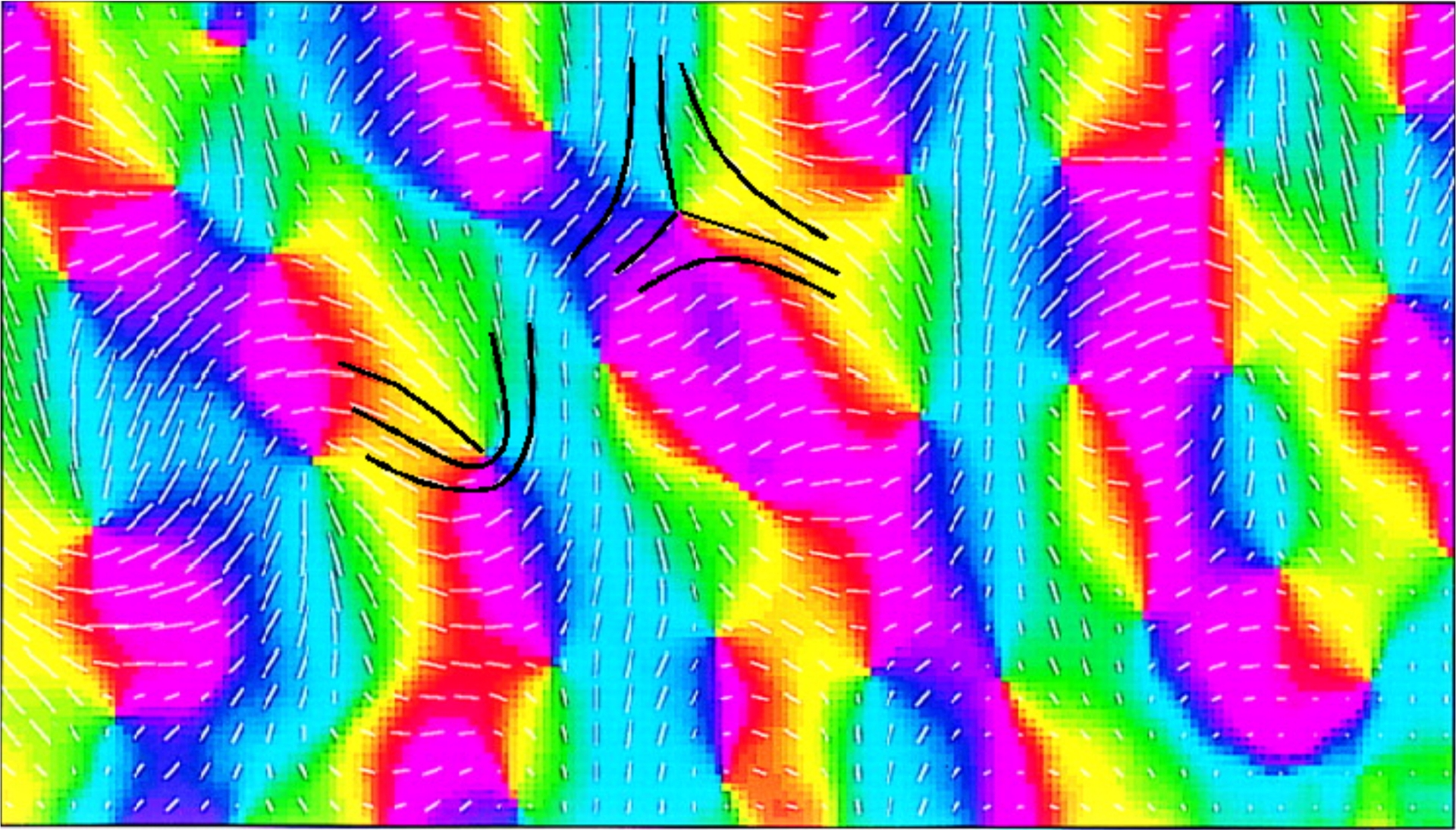
Nous allons préciser dans les Figures 12–15 la géométrie de ces lignes de champ. On y voit que les pinwheels respectivement dextrogyres et lévogyres sont associés aux deux types de singularités génériques des champs d’orientations dans le plan. Cela est dû au fait que quand le rayon tourne autour du centre du pinwheel d’un angle 𝜃, l’orientation associée tourne de 𝜃/2. Donc deux rayons diamétralement opposés correspondent à des orientations orthogonales.
La singularité « point d’arrêt » (« end point »).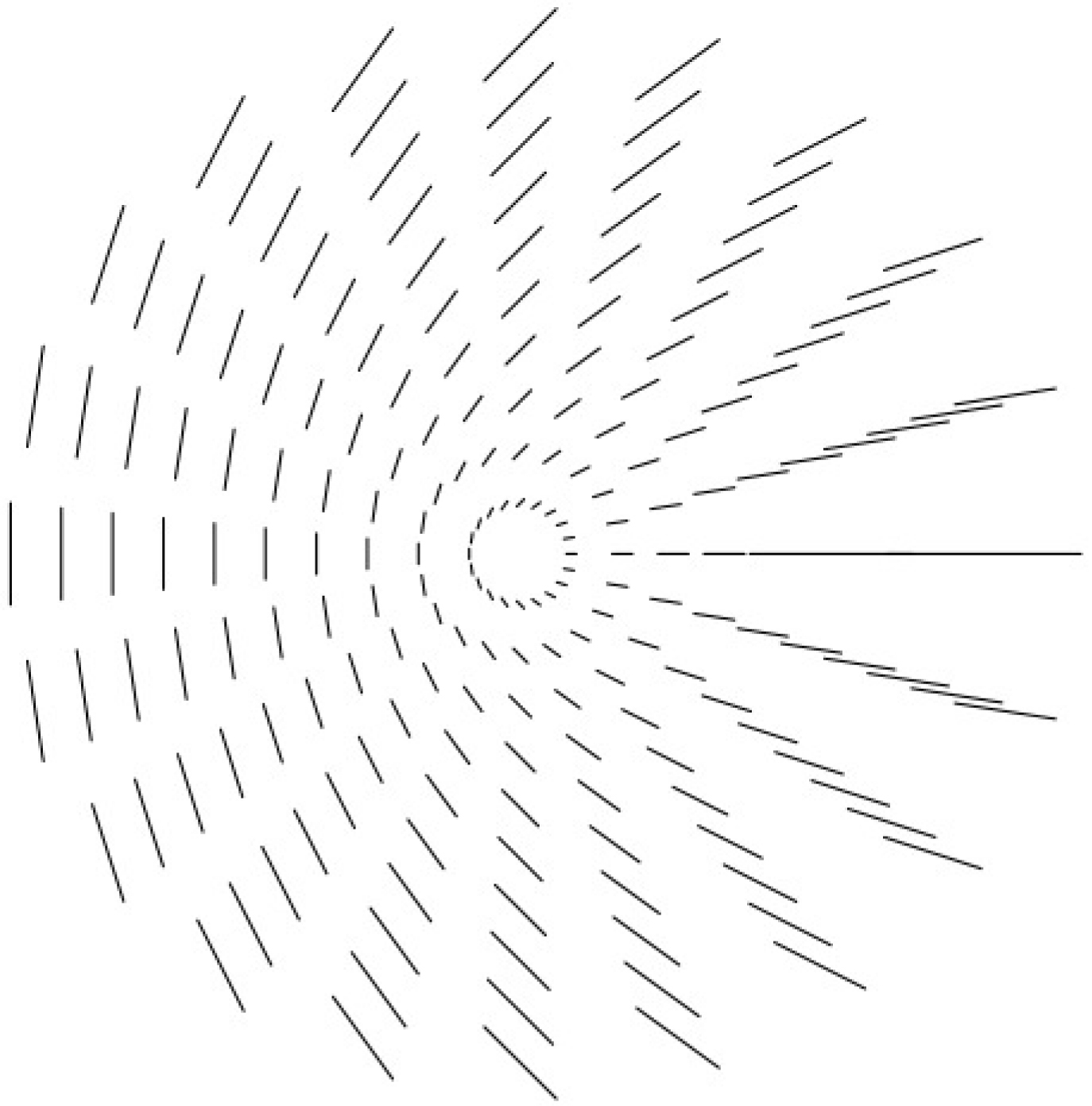
La singularité « point triple ».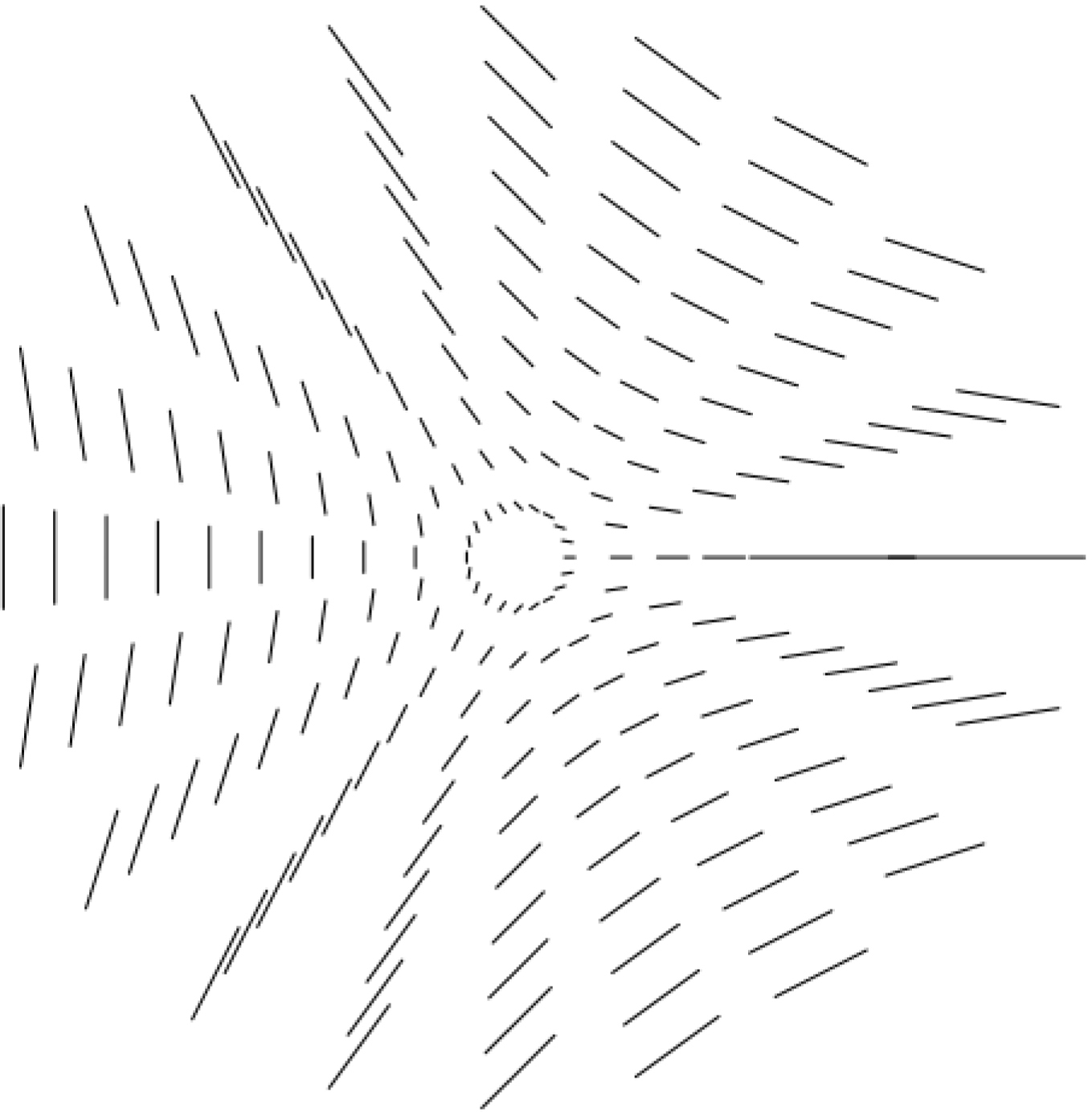
Les courbes intégrales du champ des orientations au voisinage des pinwheels. À gauche une singularité point d’arrêt, à droite une singularité point triple.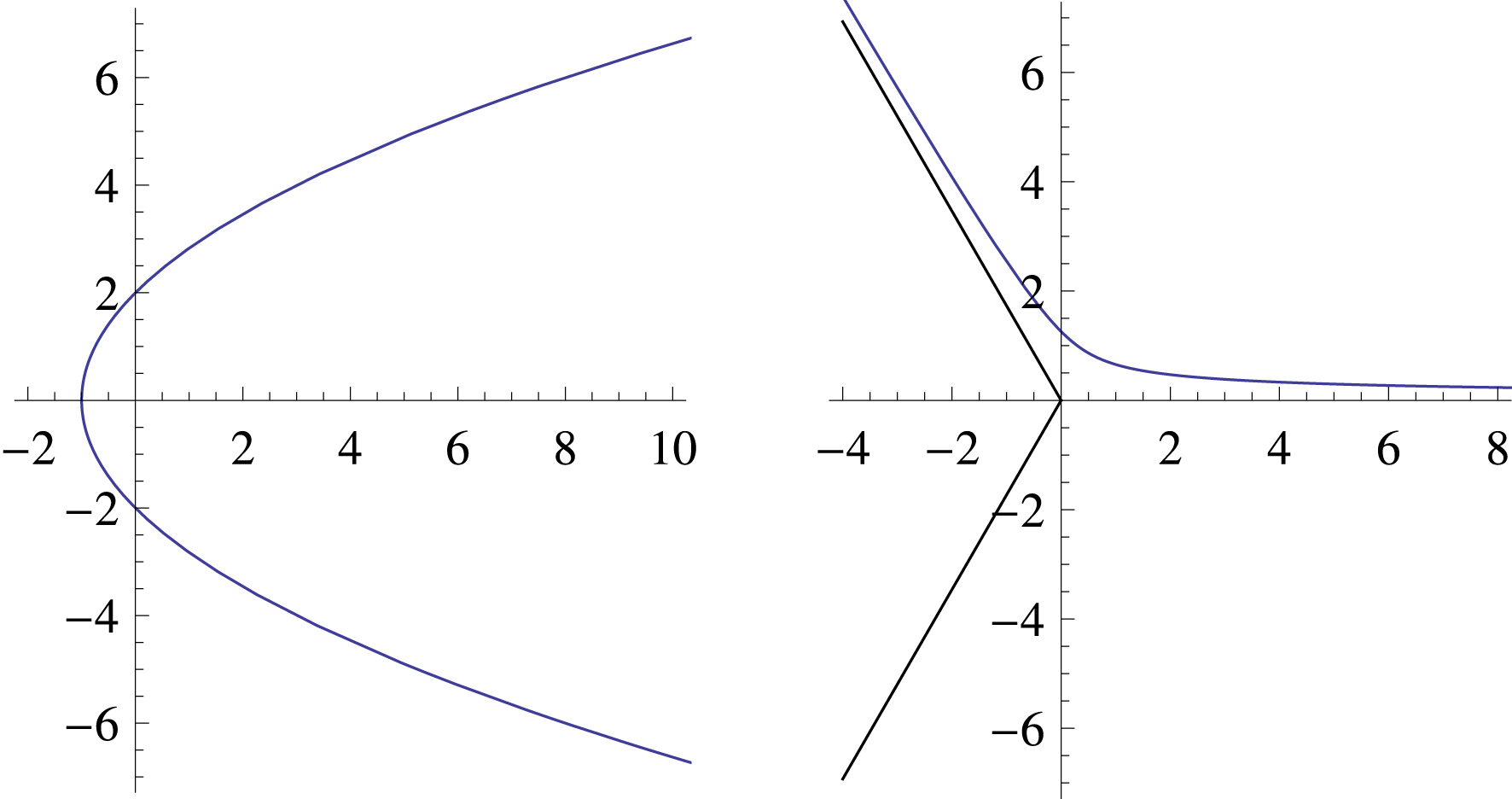
Colonnes de gauche à droite : les lignes de champ des champs 𝜑+, 𝜑−, 𝜓+, 𝜓− (voir texte) pour les quatre valeurs (de haut en bas) 0, π/8, π/4, 3π/8, du paramètre 𝛼.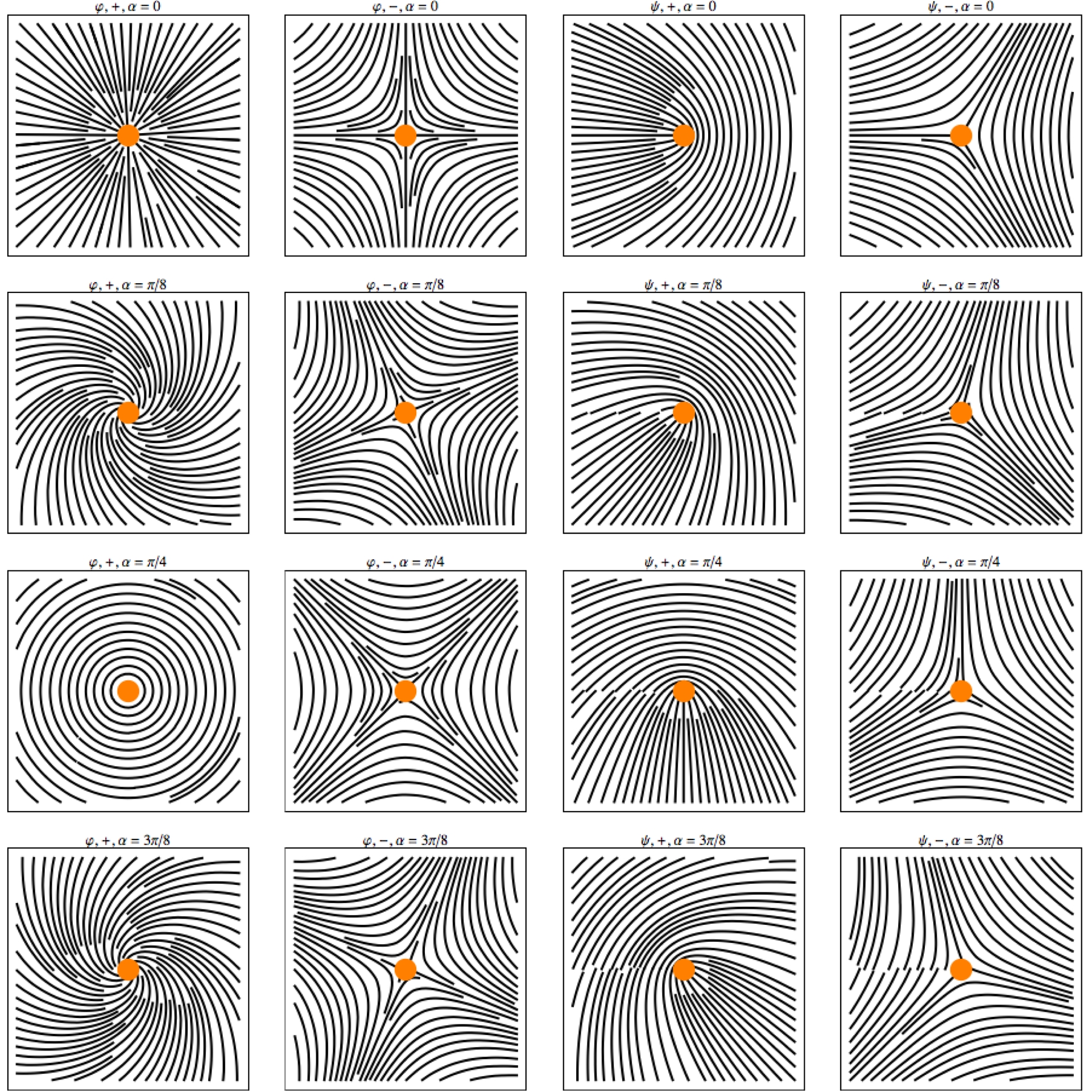
Quand on observe empiriquement les pinwheels on constate qu’il existe soit une ligne soit trois lignes où le rayon d’angle 𝜃 porte l’orientation 𝜃. Cela peut s’expliquer facilement à partir d’un angle 𝛼, caractéristique du pinwheel considéré, et qui mesure en quelque sorte l’écart entre l’orientation portée par le rayon d’angle 𝜃 du pinwheel et l’orientation 𝜃/2.
Si l’orientation 𝜓𝜃 associée avec le rayon d’angle 𝜃 est $\psi _{\theta }^{+}=\alpha +\theta /2=\varphi _{\theta }^{+}/2$, les deux directions seront les mêmes pour $\psi _{\theta _{0}}^{+}=\alpha +\theta _{0}/2=\theta _{0}$, c’est-à-dire pour 𝜃0 = 2𝛼 et alors $\varphi _{\theta }^{+}=4\alpha $. Comme 𝛼 est défini modulo π, il n’y a qu’une solution et l’on obtient le modèle local de la Figure 12 (point d’arrêt ou « end point »). Si au contraire l’orientation 𝜓𝜃 associée avec le rayon d’angle 𝜃 est $\psi _{\theta }^{-}=\alpha -\theta /2=\varphi _{\theta }^{-}/2$, les deux directions seront les mêmes pour $\psi _{\theta _{0}}^{-}=\alpha -\theta _{0}/2=\theta _{0}$, c.a.d. pour 𝜃0 = 2𝛼/3 et alors $\varphi _{\theta }^{+}=4\alpha /3$. Il y a trois solutions et l’on obtient le modèle local de la Figure 13 (point triple).
Il est très facile de calculer les courbes intégrales du champ des orientations au voisinage de ces singularités, par exemple pour 𝛼 = 0. Plaçons-nous en coordonnées polaires (𝜌,𝜃). On a x = 𝜌 cos(𝜃) et y = 𝜌 sin(𝜃) et par conséquent
| \begin{equation*} \begin{cases} \mathrm{d} x =\cos ( \theta ) \,\mathrm{d}\rho -\rho \sin ( \theta ) \,\mathrm{d}\theta\\[0.20em] \mathrm{d} y =\sin ( \theta ) \,\mathrm{d}\rho +\rho \cos ( \theta ) \,\mathrm{d}\theta. \end{cases} \end{equation*} |
| \begin {equation*} \sin ( \theta /2) \frac {\mathrm {d}\rho }{\mathrm {d}\theta }+\rho \cos ( \theta /2) =0. \end {equation*} |
| \begin {equation*} \rho =\frac {\rho _{\uppi }}{\sin ^{2}( \theta /2) }=2\frac {\rho _{\uppi }}{1-\cos ( \theta ) } \end {equation*} |
Pour les points triples, la contrainte est dy/dx = −tan(𝜃/2) et les calculs donnent les solutions
| \begin {equation*} \rho =\frac {\rho _{\uppi }}{\sin ( 3\theta /2) ^{\frac {2}{3}}}. \end {equation*} |
Lorsque 𝛼 ≠ 0, les solutions deviennent respectivement 𝜌 = 𝜌π+2𝛼/sin2(𝜃/2−𝛼) et 𝜌 = (𝜌(π+2𝛼)/3)/(sin(3𝜃/2−𝛼)2/3) et les champs d’orientations tournent de 𝛼. Mais il faut noter que les champs des 𝜑 = 2𝜓 varient plus subtilement avec 𝛼. La singularité des 𝜑+ évolue d’un nœud avec symétrie centrale (𝛼 = 0) d’abord à des foyers stables puis à un centre (𝛼 = π/4) puis à des foyers instables, alors que la singularité des 𝜑− donne différents types de cols. La Figure 15 montre les lignes de champ de 𝜑+, 𝜑−, 𝜓+, 𝜓−, pour quatre valeurs du paramètre 𝛼: 0, π/8, π/4, 3π/8.
5. Les pinwheels comme champs de phases
Nous allons maintenant étudier mathématiquement un peu plus en détail les cartes fonctionnelles d’orientation. Elles assignent, à une certaine échelle, une orientation 𝜓(a) mod π à chaque point a de la surface corticale de V 1, surface que nous modélisons pour simplifier par un plan $\mathbb{R}^{2}$ de coordonnées (x, y) identifié à un plan complexe $\mathbb{C} $ de coordonnée z. On lui associe la fonction ei𝜑 où 𝜑 est la phase 𝜑 = 2𝜓 définie mod 2π.
Ces cartes sont donc des champs de phases dont les pinwheels sont des singularités (les lignes de niveau sont les mêmes pour 𝜓 et 𝜑). On rencontre de tels champs dans de nombreux types de phénomènes physiques, en particulier dans les structures optiques et les cristaux liquides. Il existe une vaste littérature à leur sujet et, en particulier, au sujet de leurs singularités. On peut donc essayer de transférer à l’étude des cartes d’orientation un certain nombre de ces formalismes et de ces modèles.
5.1. Champs et coordonnées
Comme nous avons plusieurs espaces et plusieurs champs à considérer, fixons d’abord les notations. La couche corticale est modélisée par un plan $\mathbb{R}^{2}$. Si a = (x,y) est un point de $\mathbb{R}^{2}$, il sera souvent utile de le considérer comme l’affixe complexe z = x + iy au moyen de l’isomorphisme $\mathbb{R}$-linéaire standard de $\mathbb{C}$ avec $\mathbb{R}^{2}$. La différence entre a et z est que a est une entité géométrique alors que z est une entité numérique. Que le plan $\mathbb{R}^{2}$ avec sa structure d’espace vectoriel puisse être muni d’une structure algébrique de corps a été une grande découverte due à Argand, Gauss et Cauchy5 .
Souvent, pour étudier la structure du champ au voisinage d’un point singulier a0, il sera utile de déplacer le repère de $\mathbb{R}^{2}$ en a0 par une translation transportant 0 en a0 et une rotation appropriée des axes et de considérer alors les coordonnées polaires (𝜌,𝜃) en a0 = 0, i.e. z = 𝜌 ei𝜃, x = 𝜌 cos(𝜃), y = 𝜌 sin(𝜃). Un champ de phases assignant à chaque point $a\in \mathbb{R}^{2}$ la phase 𝜑(a) en a est donc une application $\Phi :\mathbb{R}^{2}\rightarrow \mathbb{S}^{1}$, Φ(a) = ei𝜑(a), du plan $\mathbb{R}^{2}$ dans le cercle unité $\mathbb{S}^{1}$, autrement dit une section de la fibration $\uppi :\mathbb{R}^{2}\times \mathbb{S}^{1}\rightarrow \mathbb{R}^{2}$, possédant des singularités d’un type particulier là où la phase 𝜑(a) n’est pas définie.
Supposons pour simplifier que 𝜓(a) est régulière en dehors des points singuliers où elle est indéterminée6 . Dans les modèles locaux de pinwheels de type point d’arrêt et point triple de la Section 4.3, nous avons pris 𝜓 = 𝛼 ± 𝜃/2.
Il est souvent possible et naturel d’associer à la phase d’un champ de phases une amplitude, c’est-à-dire un module r(a). Pour les cartes fonctionnelles d’orientation, Fred Wolf et Theo Geisel (cf. Wolf and Geisel, 1998; Wolf and Geisel, 2003) ont proposé d’introduire la force de la sélectivité à l’orientation préférentielle (autrement dit la largeur de la courbe d’accord, de tuning). Sous une telle hypothèse, le champ de phases Φ devient la partie « phase » d’un champ scalaire complexe $Z:\mathbb{C}\rightarrow \mathbb{C}$, z = 𝜌 ei𝜃 ↦ r(z)ei𝜑(z) (aussi noté r(a)ei𝜑(a)). En coordonnées cartésiennes, Z(a) s’écrira Z(a) = X(a) + iY (a), X et Y étant deux fonctions réelles des variables (x,y).
5.2. Les singularités d’un champ de phases
Étant donné un champ de phases Z(a) (noté suivant les contextes Z(z), X(a) + iY (a), X(z) + iY (z), r(a)ei𝜑(a) ou r(z)ei𝜑(z)), on peut analyser sa géométrie en utilisant les outils classiques de l’analyse vectorielle : gradients 𝛻, divergences, rotationnels, laplaciens Δ, etc.
Les singularités de Z sont des points où la phase 𝜑 n’est pas définie mais où Z est quant à lui bien défini mais avec une valeur particulière, 0 ou ∞. Elles correspondent aux zéros et aux pôles de Z. Si |Z| est borné, alors elles ne peuvent être que des zéros. Nous supposerons que c’est le cas. Pour un zéro, le modèle local le plus simple est ±z si Z est une fonction holomorphe, c’est-à-dire une fonction régulière de la variable complexe z qui est dérivable par rapport à z (et pas seulement par rapport à x et y séparément). Mais Z n’a a priori aucune raison d’être holomorphe. Au voisinage d’un point a0 du plan de base $\mathbb{R}^{2}$ pris pour origine 0, on a alors au premier ordre
| \begin {eqnarray*} Z( x,y) \simeq X( 0) +x\frac {\partial X}{\partial x} ( 0) +y\frac {\partial X}{\partial y}( 0) + \mathrm {i}\left ( Y( 0) +x\frac {\partial Y}{\partial x}( 0) +y\frac { \partial Y}{\partial y}( 0) \right ) \end {eqnarray*} |
| \begin {equation*} Z( a) \simeq Z( 0) +a\cdot \nabla _{0}X+\mathrm {i}a\cdot \nabla _{0}Y \end {equation*} |
| \begin {equation*} \vert Z( a) -Z( 0) \vert ^{2}\simeq R^{2}=( a\cdot \nabla _{0}X) ^{2}+( a\cdot \nabla _{0}Y) ^{2}. \end {equation*} |
Dans les situations où l’on doit analyser des champs, les singularités jouent un rôle structural déterminant et concentrent l’essentiel de l’information morphologique. Cela est dû au fait qu’en dehors des singularités les champs sont localement « rectifiables » c’est-à-dire localement trivialisable. Nous l’avons vu avec les cartes d’orientation des Figures 4–6 : en dehors des singularités que sont les centres des pinwheels, les lignes d’iso-orientation (i.e. monochromatiques) sont localement comme des droites parallèles un peu déformées.
Tout un ensemble de concepts de la théorie des singularités se révèlent donc être pertinents pour l’analyse des cartes d’orientation. Supposons le champ Z régulier en dehors des points singuliers où la phase 𝜑 est indéterminée et où Z s’annule. Comme Z = X + iY , ces points sont les intersections des courbes d’équation X = 0 et Y = 0. La condition X = 0 correspond à r cos(𝜑) = 0, i.e. 𝜑 = π/2 mod π si r ≠ 0 et Y = 0 correspond à r sin(𝜑) = 0, i.e. 𝜑 = 0 mod π si r ≠ 0. Si X = Y = 0, on a nécessairement r = 0 car les deux conditions sur 𝜑 sont incompatibles. Génériquement, deux courbes régulières dans un plan se coupent transversalement en des points isolés. C’est le cas ici des courbes X = 0 et Y = 0. Génériquement les points qui satisfont à ces deux conditions sont de codimension 2 et, comme l’espace ambiant $\mathbb{R}^{2} $ est de dimension 2, sont des points isolés7 .
5.3. Les deux champs d’orientation et d’iso-orientation
Insistons sur le fait que, comme nous l’avons indiqué plus haut, il faut bien distinguer plusieurs champs. Le champ Z est le champ des phases 𝜑(a) = 2𝜓(a). Il possède des lignes d’isophase, appelées les « fronts d’onde » par analogie avec l’optique. Comme iso-phase = iso-orientation, elles sont représentées par les lignes isochromatiques dans les cartes de pinwheels.
D’autre part, il y a le champ des orientations 𝜓(a) = 𝜑 (a)/2 interprété non plus comme une fonction angulaire sur $\mathbb{R}^{2}$ mais vraiment comme un champ définissant en chaque point a une orientation. En tant que tel, il définit un feuilletage du plan $\mathbb{R}^{2}$ par ses courbes intégrales8 . Nous avons vu à la Section 4.3 avec les modèles des points d’arrêt et des points triples quelle était la géométrie locale de ces feuilletages aux points singuliers. Notons ce champ W(a) = s(a)ei𝜓(a) en supposant que l’on ait pu attribuer une signification à l’amplitude s(a). W possède également des lignes de champ et des lignes d’isophase. Mais, contrairement à la phase 𝜑 qui est définie modulo 2π, l’angle 𝜓 n’est défini que modulo π. Pour ces pinwheels modèles, 𝜑 s’identifie en fait à 2𝛼 ± 𝜃 et 𝜓 à 𝛼 ± 𝜃 /2. On a dans ce cas ei𝜑 = (ei𝜓)2. La façon la plus simple de satisfaire cette contrainte est de prendre Z = W2.
5.4. Courant, vorticité et divergence
La différentielle d𝜑 = (∂𝜑/∂x) dx + (∂𝜑/∂y) dy correspond au vecteur gradient 𝛻𝜑 de composantes 𝛻𝜑 = (∂𝜑/∂x,∂𝜑/∂y). Le long des fronts d’onde la phase 𝜑 est constante, i.e. d𝜑 = 0, et donc, puisque (si l’on interprète comme autrefois les différentielles comme des variations infinitésimales) d𝜑 = 𝛻𝜑 ⋅da (produit scalaire), on a 𝛻𝜑 ⋅ da = 0 : les trajectoires de 𝛻𝜑 sont orthogonales aux fronts.
Pour comprendre le comportement de 𝛻𝜑 au voisinage d’une singularité, le mieux est de se placer en coordonnées polaires x = 𝜌 cos(𝜃) et y = 𝜌 sin(𝜃). Au point singulier 𝜌 = 0 le gradient 𝛻𝜑 n’est pas défini et diverge.
Pour régulariser cette situation, les physiciens ont l’habitude de considérer ce que l’on appelle le courant $\mathcal{J}$ du champ Z = rei𝜑, qui est le vecteur défini par
| \begin {equation*} \mathcal {J}=r^{2}\nabla \varphi . \end {equation*} |
| \begin {equation*} \mathcal {J}=X\nabla Y-Y\nabla X \end {equation*} |
Un autre vecteur (en fait un pseudovecteur) que considèrent les physiciens est la vorticité Ω du courant $\mathcal{J}$ c’est-à-dire, à un facteur près, son rotationnel. Par définition,
| \begin {equation*} \Omega =\tfrac {1}{2}\nabla \times \mathcal {J}=\nabla X\times \nabla Y \end {equation*} |
On remarquera que lorsque Ω = 0, soit 𝛻X = 0 ou 𝛻Y = 0 (points isolés de codimension 2), soit les gradients réels 𝛻X et 𝛻Y ont même orientation (lignes de codimension 1), autrement dit 𝛻Y = 𝛼𝛻 X pour un $\alpha \in \mathbb{R}$. La condition Ω = 0 disant que 𝛻X et 𝛻Y sont parallèles est l’opposé des conditions de Cauchy–Riemann disant que 𝛻X et 𝛻Y sont orthogonaux.
La vorticité Ω de $\mathcal{J}$ n’est pas triviale en général. Ce n’est pas le cas pour 𝛻𝜑 car le rotationnel d’un gradient 𝛻 ×𝛻 f est toujours nul en vertu du fait que pour tout vecteur on a u × u = 0. En dehors des pinwheels où il n’est pas défini, le champ 𝛻𝜑 est par conséquent irrotationnel.
Nous avons vu plus haut que, au premier ordre au voisinage d’un point a0 pris comme origine 0, le module de Z est donné par
| \begin {equation*} \vert Z( a) -Z( 0) \vert ^{2}=R^{2}=( a\cdot \nabla _{0}X) ^{2}+( a\cdot \nabla _{0}Y) ^{2}. \end {equation*} |
| \begin {eqnarray*} \mathcal {J}( a) &=&\mathcal {J}( x,y) \\ &\simeq & \left ( \begin {array}{@{}c@{}} \displaystyle y\left ( \frac {\partial X}{\partial y}( 0) \frac {\partial Y}{ \partial x}( 0) -\frac {\partial X}{\partial x}( 0) \frac {\partial Y}{\partial y}( 0) \right ) ,\\[1em] \displaystyle x\left ( -\frac {\partial X}{\partial y}( 0) \frac {\partial Y}{ \partial x}( 0) +\frac {\partial X}{\partial x}( 0) \frac {\partial Y}{\partial y}( 0) \right ) \end {array} \right ) \\ &=&( a\cdot \nabla _{0}X) \nabla _{0}Y-( a\cdot \nabla _{0}Y) \nabla _{0}X\\ &=&\Omega _{0}\times a=\omega _{0}( -y,x). \end {eqnarray*} |
Cela permet d’évaluer $|\mathcal{J}|=r^{2}|\nabla \varphi |\approx |\omega |\rho $ au voisinage des pinwheels où 𝜔 ≠ 0. Mais localement 𝜑 est constante sur les rayons d’un tel point singulier et 𝛻𝜑 est orthogonal aux rayons et donc, puisqu’en coordonnées polaires 𝛻𝜑 = (∂𝜑/∂𝜌)e𝜌 + (1/𝜌)(∂𝜑/∂𝜃)e𝜃 (avec e𝜌 le vecteur unitaire du rayon en a et e𝜃 le vecteur unitaire orthogonal à e𝜌, i.e. e𝜌 tourné de + π/2), on a 𝛻𝜑 ≃ (1/𝜌)(∂𝜑/∂𝜃)e𝜃. D’où la formule
| \begin {equation*} r^{2}\left \vert \frac {\partial \varphi }{\partial \theta }\right \vert \approx \rho ^{2}\vert \omega \vert . \end {equation*} |
« equal area vectors of the core anisotropy ellipse [r2 = cst] are swept out in equal intervals of phase. » (p.41)
Remarquons que l’excentricité des ellipses mesure l’anisotropie de la vorticité. Comme nous l’avons vu, il n’y a isotropie (les ellipses ne sont des cercles) que si les conditions de Cauchy-Riemann sont satisfaites.
5.5. Équation de Helmholtz
Nous avons vu en présentant les données expérimentales sur les pinwheels que ceux-ci apparaissent lorsqu’on superpose les différentes cartes de réponse aux différentes orientations. Nous avons vu également qu’il existe une maille caractéristique du réseau de pinwheels (cf. Section 4.1). Ces deux faits empiriques suggèrent de considérer le champ Z comme une superposition de champs plus simples présentant une longueur caractéristique. Par ailleurs, sur le plan mathématique tout champ peut être considéré à travers sa transformée de Fourier comme une superposition d’ondes planes. Les ondes planes sont les plus simples des champs possédant une longueur caractéristique. Elles s’écrivent Aei𝜅⋅a où A est une amplitude complexe Eei𝜙 et 𝜅 = (𝜅x,𝜅y) un vecteur appelé vecteur d’onde dont le module k = |𝜅|, appelé nombre d’onde, est l’analogue d’une quantité de mouvement et est associé à la longueur d’onde Λ = 2π/k (plus la longueur d’onde est petite plus le nombre d’onde est grand). Lorsqu’elles évoluent dans le temps, leur prototype est Aei(𝜅⋅a−𝜔t) où 𝜔 est une fréquence angulaire (ou pulsation) associée à la fréquence 𝜈 = 𝜔 /2π et à la période T = 1/𝜈 = 2π/𝜔.
La question de savoir si des décompositions mathématiques d’entités d’un certain type (ici des champs) en superpositions d’entités de même type mais élémentaires (ici des ondes planes) sont des décompositions « réelles » ayant un sens physique, est un vieux problème épistémologique qui remonte à l’antiquité. Déjà quand Ptolémée décrivait les trajectoires « errantes » des planètes comme compositions de mouvements circulaires uniformes sur des épicycles9 , les « physiciens » critiquaient les « mathématiciens » en arguant que seules les trajectoires observées avaient une réalité physique et que les épicycles n’étaient que des artéfacts mathématiques. Mais en fait il faut bien comprendre que de telles décompositions servent à calculer et qu’elles sont indispensables pour reconstruire computationnellement les données empiriques. De telles « synthèses computationnelles » n’ont pas de contenu ontologique. Elles sont justement « computationnelles ». C’est le cas ici avec les décompositions en ondes planes.
Les superpositions d’onde planes ont été utilisées entre autres par Theo Geisel, Matthias Kaschube, Michael Schnabel1 et Fred Wolf (cf. par exemple Kaschube et al., 2008) pour construire de remarquables modèles d’auto-organisation et d’évolution par bifurcations successives de cartes d’orientations lors d’un processus d’apprentissage. Dans ce qui suit nous supposerons que la carte d’orientation est stabilisée.
Il est trivial de vérifier que les ondes planes U = Aei𝜅⋅a satisfont une équation fondamentale, dite équation de Helmholtz, ΔU + k2 U = 0. En effet, 𝜅 ⋅ a = x𝜅x + y𝜅y et donc
| \begin {equation*} \Delta U=\frac {\partial ^{2}U}{\partial x^{2}}+\frac {\partial ^{2}U}{ \partial y^{2}}=-A\kappa _{x}^{2}\mathrm {e}^{\mathrm {i}\kappa \cdot a}-A\kappa _{y}^{2}\mathrm {e}^{\mathrm {i}\kappa \cdot a}=-k^{2}U. \end {equation*} |
| \begin {equation*} \Delta Z+k^{2}Z=0. \end {equation*} |
Il est intéressant d’illustrer un exemple. Nous le ferons en suivant une analogie avec les champs de phases en optique.
6. Illustration d’un exemple
6.1. La structure du champ
La Figure 16, inspirée d’un travail de Berry (2009) sur les courants optiques montre une superposition de 10 ondes planes de même k. On voit à quel point ce champ de phases ressemble à nos pinwheels, avec ses lignes d’isophase (i.e. d’iso-orientation), ses lignes de gradient orthogonales et ses points cols.
La structure du champ de phases Z qui est une superposition de 10 ondes planes de même nombre d’onde k. Les vecteurs d’ondes 𝜅 sont donnés dans la Table 1 tirée de Berry (2009). Les lignes d’isophase sont en orange et les lignes de courant en bleu.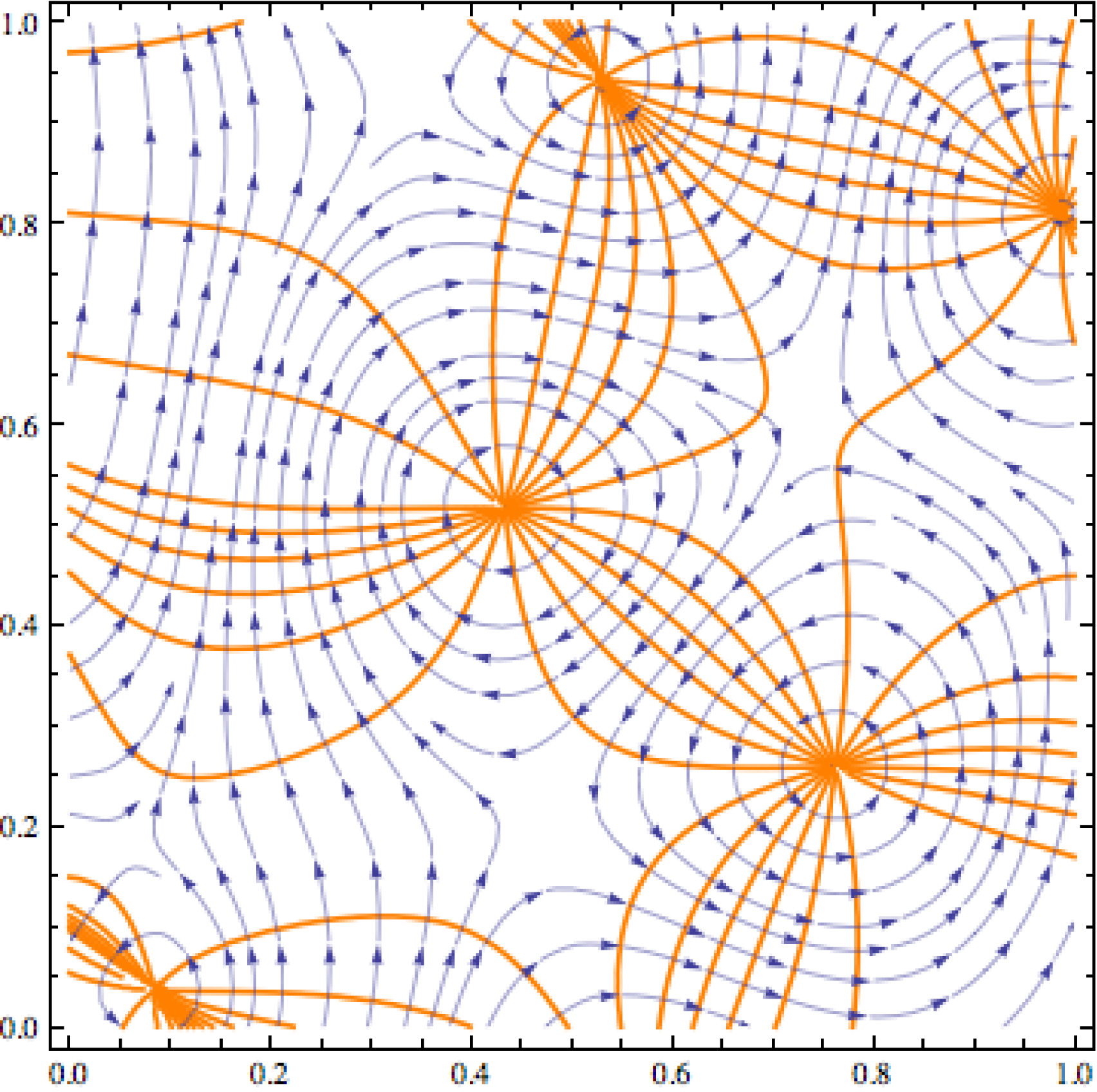
Plus précisément, Z est une superposition de 10 ondes planes
| \begin {eqnarray*} Z&=&\sum _{j=1}^{j=10}E_{j}\exp ( \mathrm {i}( \phi _{j}+2\uppi x\cos ( \alpha _{j}) +2\uppi y\sin ( \alpha _{j}) ) ) \\ &=&r\mathrm {e}^{\mathrm {i}\varphi } \end {eqnarray*} |
Les 𝛼j, 𝜙j, Ej des 10 ondes planes dont la superposition est montrée la Figure 16
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | |
|---|---|---|---|---|---|---|---|---|---|---|
| 𝛼j | 5.971 | 2.666 | 0.939 | 4.629 | 1.023 | 1.537 | 2.710 | 3.273 | 4.356 | 5.032 |
| 𝜙j | 3.846 | 0.777 | 5.008 | 2.916 | 6.274 | 4.344 | 2.411 | 5.688 | 1.734 | 0.214 |
| Ej | 0.337 | 0.015 | 0.762 | 0.785 | 0.625 | 0.442 | 0.688 | 0.065 | 0.064 | 0.035 |
Si l’on note Φj les phases 𝜙j + 2πx cos(𝛼j) + 2πy sin(𝛼j), on obtient une expression $Z=\sum _{j=1}^{j=10}E_{j}\exp ( \mathrm{i}\Phi _{j}) $ sur laquelle il est facile de calculer. La Figure 17 montre les pinwheels de Z. Les lignes blanches sont des coupures où 𝜑 saute de 2π, cela étant dû au fait que 𝜑 est à valeurs dans $\mathbb{S}^{1}$ mais est représenté comme ayant ses valeurs dans $\mathbb{R}$. Les coordonnées des 5 pinwheels sont {0.528545, 0.942654}, {0.988124, 0.811337}, {0.433271, 0.516137}, {0.761954, 0.258734}, {0.0838329, 0.0359263}. Ils sont donnés par les intersections des lignes de niveau X = 0, Y = 0. La Figure 18 représente ces lignes, X = 0 en rouge et Y = 0 en bleu.
Pinwheels du champ de phases Z pour x,y ∈ [0,1]. Les lignes blanches sont des coupures où 𝜑 saute de 2π.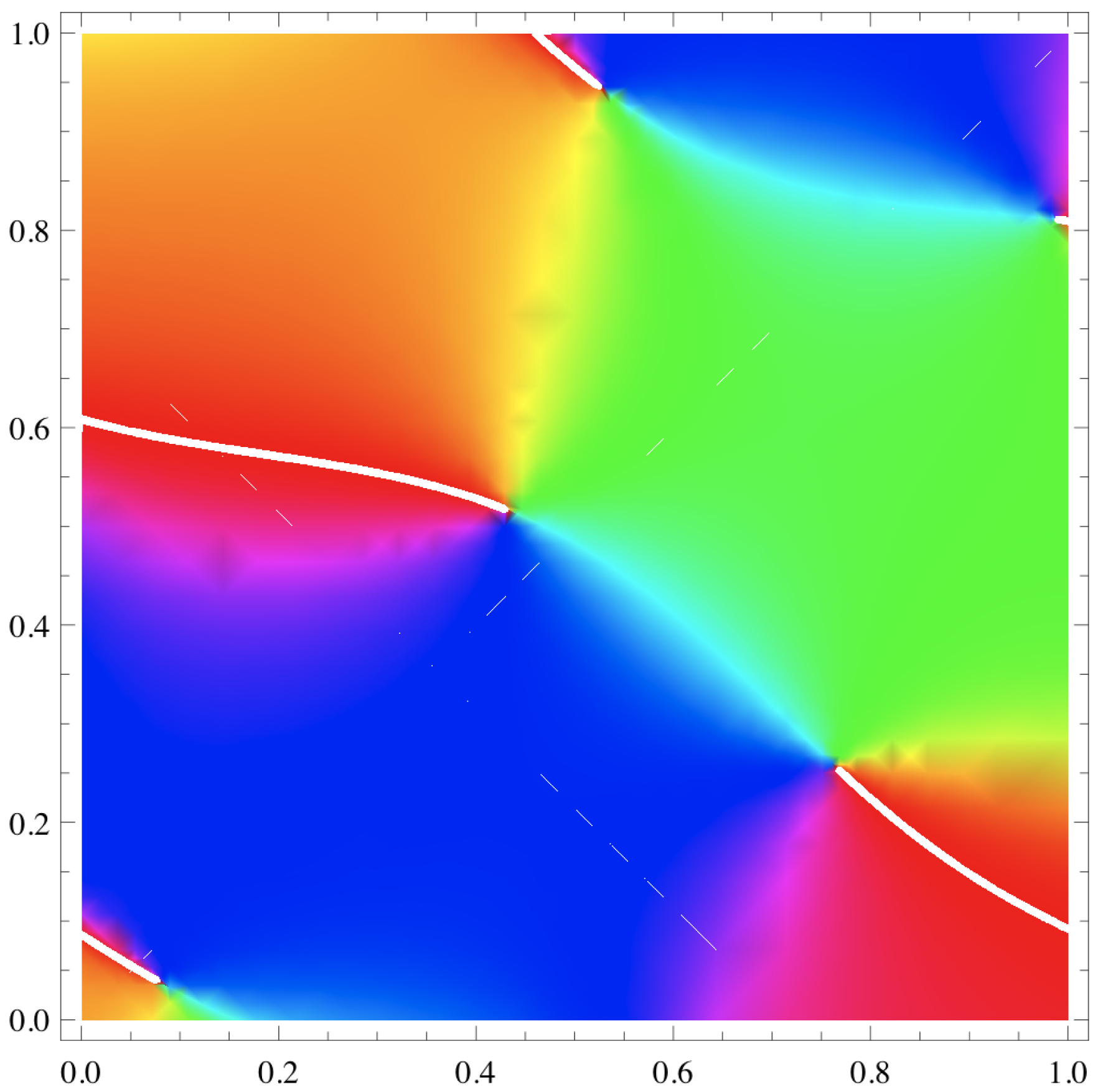
Lignes X = 0 (en rouge) et Y = 0 (en bleu) du champ de phases Z.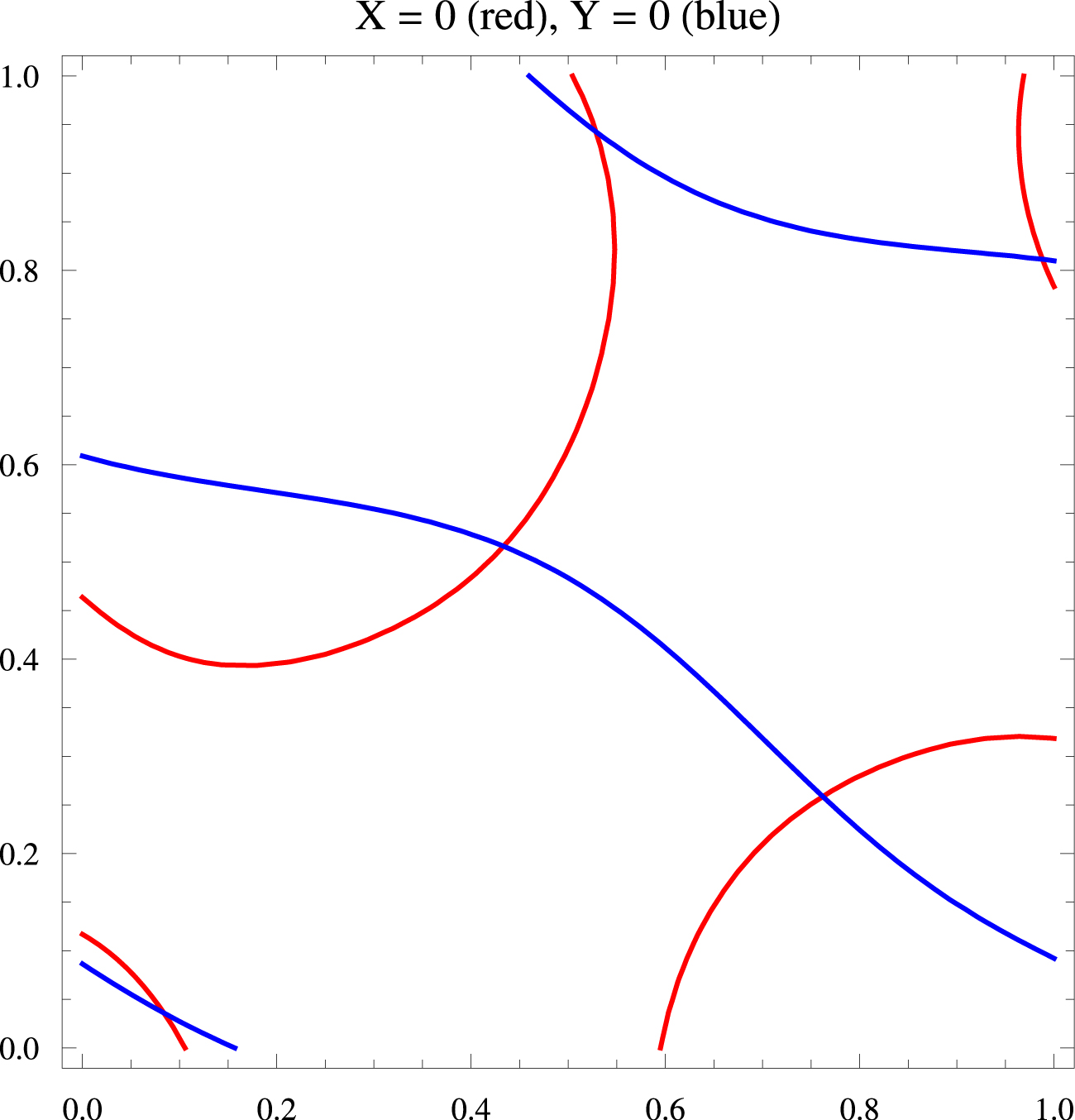
La Figure 19 montre quant à elle la structure du champ de phases Z sur le carré x,y ∈ [0,1]. La première ligne montre le module r de Z
Structure du champ de phases Z pour x,y ∈ [0,1]. Première ligne: module r de Z; (i) lignes de niveau; (ii) fonction r(x,y) sous deux perspectives, la seconde permettant de voir les points singuliers (dislocations) où r = 0 ainsi que les maxima de r. Seconde ligne: argument 𝜑 de Z; (i) lignes de niveau; (ii) fonction 𝜑(x,y) sous deux perspectives, la seconde permettant de voir les coupures où 𝜑 saute de 2π.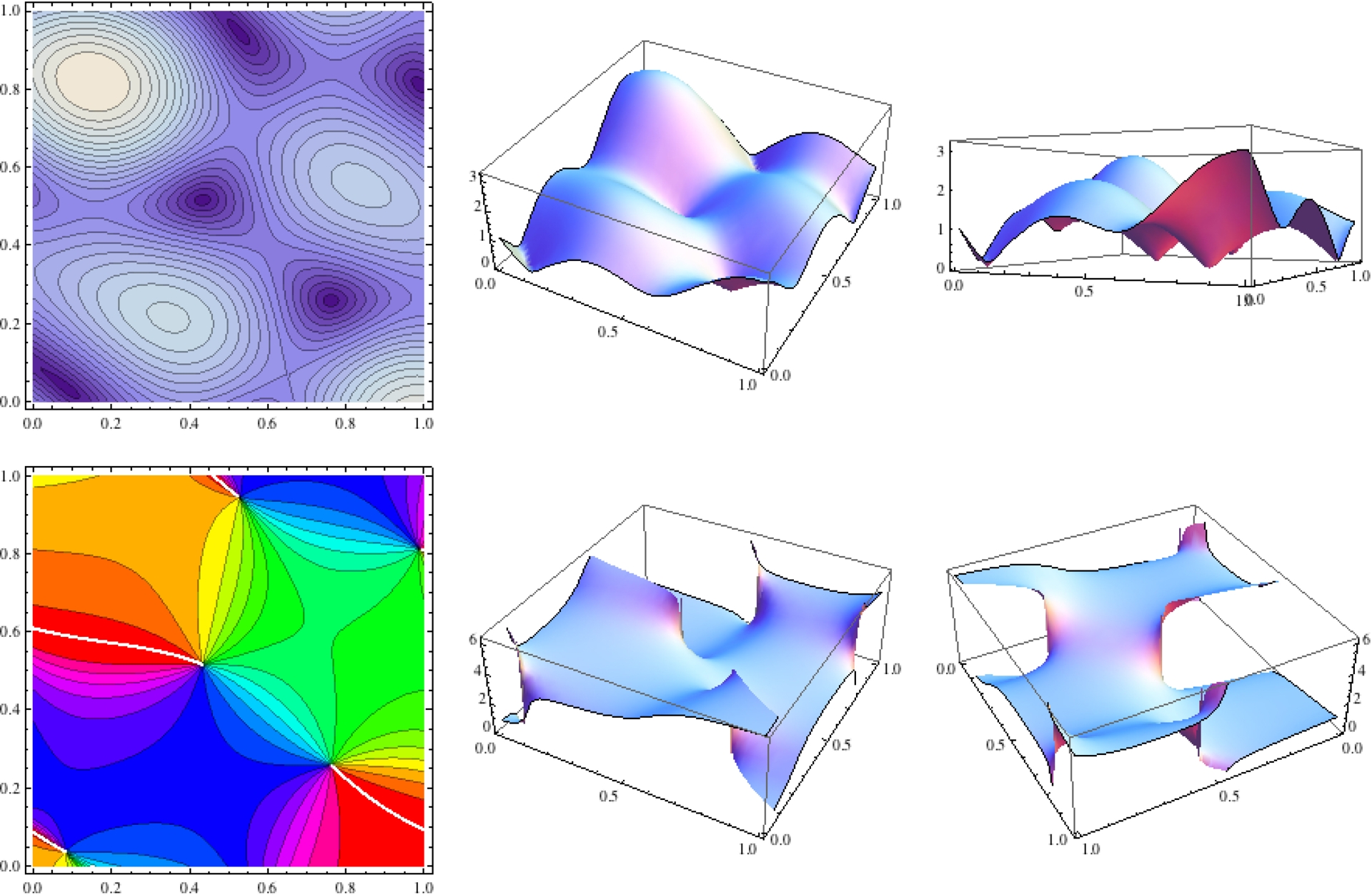
- d’abord avec ses lignes de niveau puis
- en tant que fonction r(a) = r(x,y) sous deux perspectives, la seconde permettant de bien voir les points singuliers (dislocations) où r = 0 et les maxima de r.
La seconde ligne montre l’argument 𝜑 de Z (i) d’abord avec ses lignes de niveau (on retrouve celles de la Figure 16) puis (ii) en tant que fonction 𝜑(a) = 𝜑 (x,y) sous deux perspectives, la seconde permettant de bien voir les coupures où 𝜑 saute de 2π.
La Figure 20 montre plus de pinwheels de Z (x,y ∈ [0,3]), les coupures blanches y représentant 𝜑 = 0 = 2π. On notera qu’il y a 29 pinwheels dans une aire de 32 = 9, ce qui donne une densité d de 29/9 ∼ 3.2. Nous expliquerons plus bas une formule donnant d = π /Λ2, soit, dans notre cas où Λ = 1, d = π. On voit que l’approximation est excellente.
Pinwheels de Z pour x,y ∈ [0,3]. Les coupures blanches représentent 𝜑 = 0 = 2π.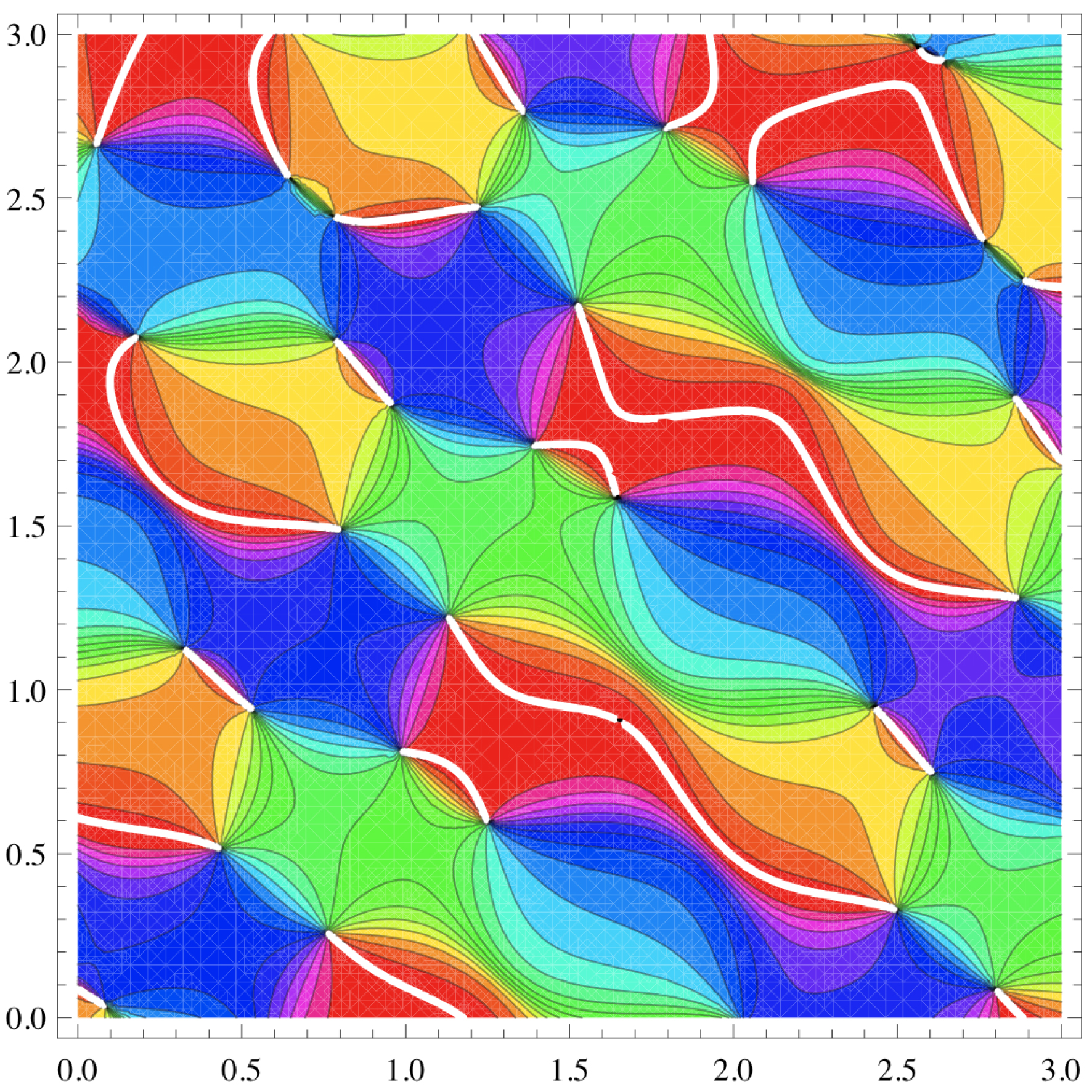
Nous représentons également à la Figure 21 les lignes d’orientation, c’est-à-dire les lignes de champ de W, ainsi que les lignes de champ de Z (à ne pas confondre avec les lignes d’isophase). On retrouve bien des occurrences des singularités typiques illustrées à la Figure 15 de la Section 4.3. À gauche nous voyons un point triple et un point d’arrêt comme dans les deux colonnes de droite de la Figure 15 et à droite un point col et un point foyer comme dans les deux colonnes de gauche de cette figure.
Lignes de champ de W et de Z dans le cas de l’exemple de la Figure 19. À gauche un point triple et un point d’arrêt comme dans les deux colonnes de droite de la Figure 15 et à droite un point col et un point foyer comme dans les deux colonnes de gauche de cette figure.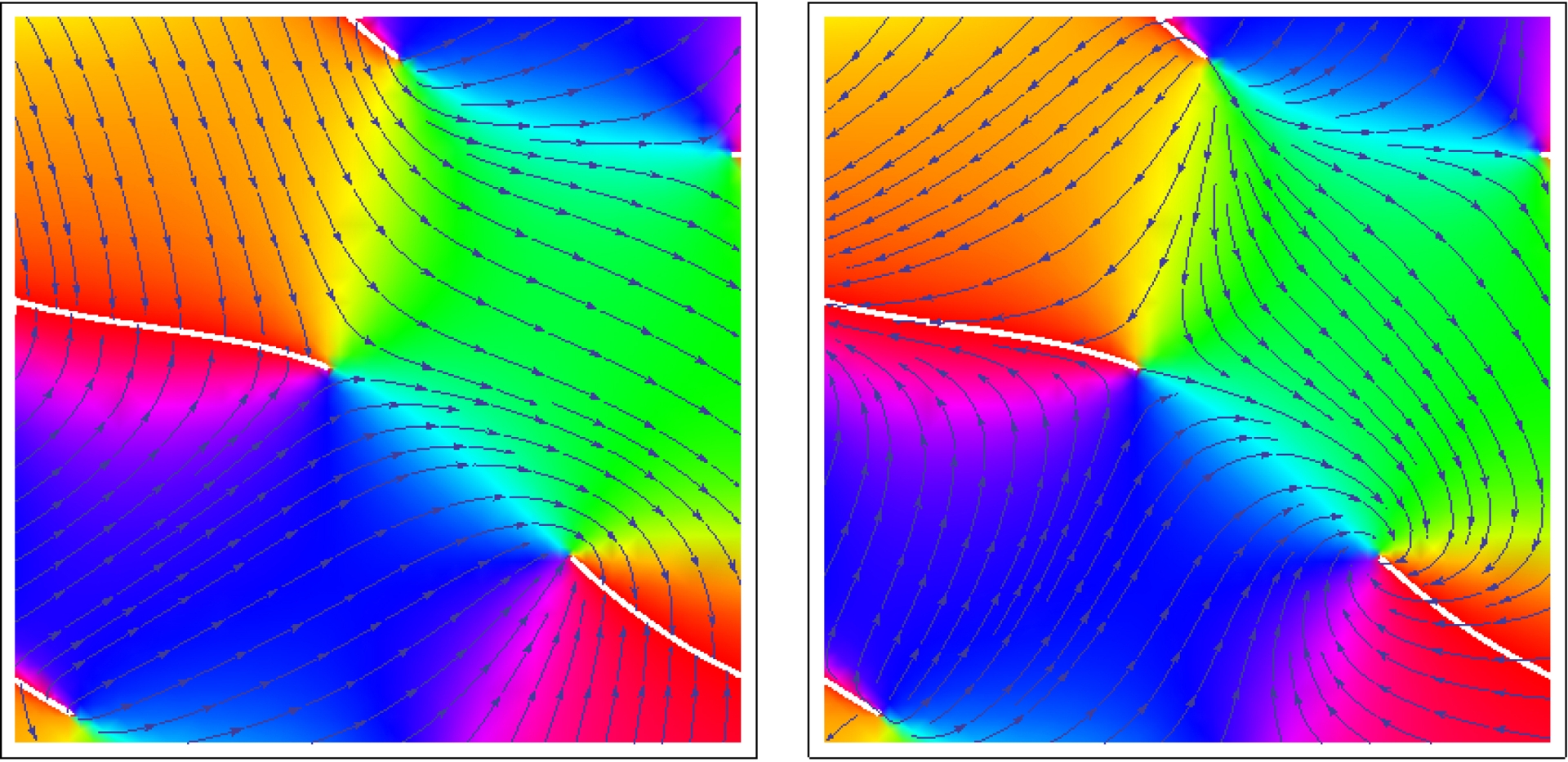
6.2. Statistique des pinwheels comme singularités de phase
Les cartes des pinwheels comme champs de phases peuvent être très variées. Il est donc intéressant d’en faire une étude statistique à partir de certaines hypothèses simplificatrices. De telles études ont déjà été menées à bien en optique en particulier par Michael Berry et Mark Richard Dennis (cf. par exemple Berry and Dennis, 2000; Berry and Dennis, 2007). Il s’agit d’un sujet où ont convergé des études de Wolf, Geisel et Kaschube (cf. Wolf and Geisel, 1998; Wolf and Geisel, 2003; Kaschube et al., 2008)10 , celles d’un groupe de travail autour de Daniel Bennequin, et également des travaux de Citti, Sarti et d’un de leurs doctorants Barbieri et al. (2012).
Dans sa thèse (Dennis, 2001), Dennis donne des résultats précis pour les superpositions d’ondes planes
| \begin {equation*} Z=\sum _{\kappa }A_{\kappa }\mathrm {e}^{\mathrm {i}\kappa \cdot a} \end {equation*} |
Une simplification supplémentaire consiste à considérer des ondes « monochromatiques » de même nombre d’onde k, le vecteur d’onde 𝜅 variant donc sur le cercle de rayon k. Dans ce cas, $\mathcal{R}( u) $ devient le Dirac 𝛿(u − k). Cette hypothèse correspond au fait que Z est solution de l’équation de Helmholtz.
On peut alors calculer la densité moyenne d des dislocations de phase. Comme celles-ci sont définies par les conditions X = 0, Y = 0, elle sera donnée par la moyenne de 𝛿(X)𝛿(Y ) relativement à la mesure dX dY . Relativement à la mesure dx dy il faut faire intervenir le jacobien de Z(x,y) = X(x,y) + iY (x,y), c’est-à dire
| \begin {equation*} \vert \omega \vert =\vert \nabla X\wedge \nabla X\vert =\left \vert \frac {\partial X}{\partial x}\frac {\partial Y}{\partial y}- \frac {\partial X}{\partial y}\frac {\partial Y}{\partial x}\right \vert . \end {equation*} |
| \begin {eqnarray*} d=K/4\uppi \ \mathrm {avec}\ K=\int _{0}^{\infty }k^{2}\mathcal {R} ( k)\,\mathrm {d}k=\langle k^{2}\rangle _{\mathcal {R}} \end {eqnarray*} |
« the results apply to any complex scalar random function. » (…) « The geometry thus revealed is extraordinary complicated and occasionally counterintuitive. » (p. 2076)
Le nombre d’onde k est proportionnel à l’inverse d’une longueur d’onde Λ = 2π/k et donc Λ2 = 4π2/k2 et k2/4π = π /Λ2. Par conséquent la densité de singularités d est la moyenne $\langle \uppi /\Lambda ^{2}\rangle _{\mathcal{R}} $. Le d = π/Λ2 de la section précédente 6.1 en est un cas particulier.
Nous avons vu à la Figure 20 qu’il y avait 29 pinwheels dans une aire de 32 = 9, ce qui donnait une densité empirique de d de 29/9 ∼ 3.2 pour une densité théorique de d = π puisque Λ = 1. Le résultat est remarquable.
7. Conclusion
Dans cet article nous avons surtout discuté la façon dont les cartes d’orientations implémentent (à travers un collapse dimensionnel 2D⟶3D) une version discrétisée de la fibration π : R × P→R ayant pour base l’espace rétinien R et pour fibre la droite projective des orientations P. Nous n’avons rien dit des connexions « horizontales » cortico-corticales à longue portée qui connectent des éléments de contact (a,p) appartenant à des hypercolonnes différentes. Elles sont pourtant fondamentales car elles sont extrèmement anisotropes et connectent entre eux des éléments de contact (a1,p1) et (a2,p2) tels que (i) p1 et p2 soient approximativement parallèles11 et (ii) la direction commune p soit celle de la droite (a1,a2). Comme l’explique William Bosking dans (Bosking et al., 1997) :
« The system of long-range horizontal connections can be summarized as preferentially linking neurons with co-oriented, co-axially aligned receptive fields ».
Cette architecture très spécifique hautement anisotrope explique la façon dont le système visuel primaire est capable d’intégrer des données locales en contours globaux et même de construire des contours illusoires qui complètent des données sensorielles lacunaires. Elle est à la base des propriétés gestaltistes, bien connues mais bien énigmatiques, de la perception. Elle est extrêmement intéressante à mathémaiser et nous lui avons consacré de nombreuses et longues études. Le lecteur intéressé pourra consulter par exemple Petitot (1999; 2003; 2006) ou Petitot (2014).
Une fois modélisée à un niveau mésoscopique les deux grandes classes de connexions internes à V 1 dont nous avons parlé d’emblée dans notre introduction, celle des connexions « verticales » intra-columnaires isotropes et à courte portée et celle des connexions « horizontales » cortico-corticales inter-columnaires anisotropes et à longue portée, la modélisation des niveaux micro sous-jacents devient mieux contrôlable sur le plan théorique. Elle peut échapper aux limites du « tout computationnel » anti-conceptuel. Comme en optique, la mésogéométrie commande la microphysique.
En optique, il existe trois niveaux : géométrique, ondulatoire, quantique. Dans l’analogie, le niveau géométrique correspond à notre niveau méso-géométrique. Le niveau ondulatoire correspond au traitement que nous venons d’effectuer en identifiant les pinwheels à des singularités de champs de phases. Mais, ainsi que le note Berry (2009), l’optique ondulatoire est une moyenne sur des interactions microphysiques relevant de l’optique quantique et
« the current gives the time-averaged force on small particules ».
Le gradient de phase 𝛻𝜑 donne le moment induit sur les particules par les impacts des photons individuels. Et comme la probabilité de ces impacts est r2, le moment moyen est bien le courant $\mathcal{J}=r^{2}\nabla \varphi $.
On peut ainsi faire l’hypothèse qu’il existe une microphysique d’événements élémentaires dont la mésogéométrie des cartes d’orientation est une sorte de squelette morphologique. Ce sont sans doute les potentiels d’action qui jouent le rôle de « petites particules ».
Remerciements
Je remercie Olivier Faugeras de m’avoir proposé de participer à ce numéro thématique ainsi que Madame Isabelle Vallet pour son aide éditoriale. Je remercie aussi beaucoup le reviewer pour ses remarques constructives.
Declaration of interests
L’auteur ne travaille pas, ne conseille pas, ne possède pas de parts, ne reçoit pas de fonds d’une organisation qui pourrait tirer profit de cet article, et n’a déclaré aucune autre affiliation que son organisme de recherche.
1 La traduction anglaise de certaines sources de cet article a été intégrée à mon ouvrage (Petitot, 2017).
2 Dans cet article, l’auteur compare le macaque (primate, diurne, yeux frontaux), le chat (carnivore, crépusculaire, yeux frontaux), le tree shrew ou tupaya (scandentia, diurne, yeux latéraux), l’écureuil gris ou gray squirrel (rongeur, diurne, yeux latéraux), le rat (rongeur, nocturne, yeux latéraux) et le singe de nuit, singe hibou, douroucoulis ou Owl Monkey (primate du Nouveau Monde, nocturne, yeux frontaux), le furet, etc. Plus récemment (2022), Jung et al. (2022) ont traité le cas de marsupiaux australiens.
3 Les géomètres parlent de directions là où les neurophysiologistes parlent d’orientations.
4 Dans la modélisation géométrique des architectures fonctionnelles neurales, on rencontre de nombreux conflits terminologiques. Des items lexicaux comme « fibre », « projection », « connexion », etc. sont utilisés avec des sens différents par les mathématiciens et les neurophysiologistes. Le contexte permet en général de lever facilement les ambiguïtés.
5 Un théorème profond dit que les seuls $\mathbb{R}^{n}$ que l’on peut munir d’une structure de corps sont $\mathbb{R}$ (le corps des nombres réels), $\mathbb{R}^{2}$ (le corps des nombres complexes), $\mathbb{R}^{4}$ (le corps non commutatif des quaternions de Hamilton) et $\mathbb{R}^{8}$ (le corps non associatif et non commutatif des octonions de Cayley). Cela est lié au fait que les seules sphères $\mathbb{S}^{n-1}$ « parallélisables » (i.e. dont le fibré tangent est isomorphe au produit direct $\mathbb{S}^{n-1}\times \mathbb{R}^{n-1}$) sont $\mathbb{S}^{1}$, $\mathbb{S}^{3}$ et $\mathbb{S}^{7} $ (Frank Adams). $\mathbb{R}^{3}$ n’a pas de structure de corps et $\mathbb{S}^{2}$ n’est pas parallélisable : tout champ de vecteurs sur $\mathbb{S}^{2}$ s’annulle quelque part (théorème de Poincaré–Hopf, quand on « peigne » une $\mathbb{S}^{2}$ « chevelue » il y a toujours un « épi » quelque part).
6 Dans certains cas il peut y avoir, en plus des pinwheels, des « fractures » dans les cartes d’orientation.
7 Dans la théorie des mésophases (cristaux liquides), ces singularités de phase sont appelées des « dislocations ».
8 Un feuilletage du plan est une décomposition en une famille de courbes.
9 Le titre « Hê Megálê Súntaxis » de son célèbre traité l’« Almageste » signifie précisément « La Grande Composition ».
10 Dans ces articles, la statistique des pinwheels est calculée en détail dans le cadre des modèles d’auto-organisation et d’évolution des cartes d’orientations.
11 Le transport parallèle est l’un des principaux fondements de la géométrie comme l’ont bien expliqué Élie Cartan et Hermann Weyl.