



 CC-BY 4.0
CC-BY 4.0
1. Introduction
Les activités anthropiques augmentent la concentration de dioxyde de carbone (CO2), avec une hausse documentée de 340 à 415 ppm entre 1980 et 2020, et des projections qui varient entre 540 et 1300 ppm d’ici la fin du siècle. Cette augmentation du CO2 s’accompagne d’une augmentation des températures moyennes, qui pourrait atteindre +4 °C d’ici 2100 (Calvin et al., 2023). Les évènements climatiques extrêmes, tels que les vagues de chaleur et les sécheresses, sont susceptibles de gagner en fréquence et en intensité dans de nombreuses zones agricoles, ce qui représente une menace croissante pour les écosystèmes naturels et agricoles (Zscheischler et al., 2018). Les plantes étant des organismes sessiles, la plasticité phénotypique, définie ici comme la variation phénotypique des caractères en réponse à l’environnement (Hallgrimsson et al., 2019) et aux pratiques culturales, est cruciale pour leur adaptation à des conditions environnementales variables, telles que celles engendrées par le changement climatique.
La plasticité des plantes face à un seul facteur environnemental a été largement étudiée pour les niveaux élevés de CO2 (eCO2 ; par exemple Ainsworth and Long, 2021), les températures élevées (c’est-à-dire une augmentation de la température quotidienne moyenne), les vagues de chaleur (par exemple, Farooq et al., 2011) et le déficit hydrique (par exemple, Vadez et al., 2024). Lorsque les facteurs quantitatifs de croissance (CO2, eau) et les conditions de croissance (température, eau) varient en dehors de leur plage optimale pour les plantes, ils induisent des stress (déficit hydrique ou excès d’eau, chaleur ou froid), définis ici comme un « changement dans l’environnement susceptible de réduire ou de modifier négativement la croissance ou le développement des plantes » de façon irréversible (Levitt, 1972; Salisbury and Ross, 1992). Les stress thermiques résultant de températures suboptimales, froides ou élevées, pendant la journée ou la nuit, et les déficits hydriques édaphiques ou atmosphériques (c.-à-d. dans le sol et/ou dans l’atmosphère) constituent les stress abiotiques majeurs limitant la croissance des plantes. Toutefois, dans un contexte de changement climatique, ces différents facteurs environnementaux sont amenés à varier conjointement, les interactions en jeu étant encore mal comprises ; ces interactions ont été particulièrement étudiées aux échelles moléculaires, montrant dans de nombreux cas une signature unique de stress combinés (Zandalinas and Mittler, 2022), mais ont besoin d’être mieux appréhendées aux échelles de la plante et du peuplement pour prédire les rendements futurs et les meilleures stratégies d’adaptations (Heino et al., 2023; Lobell and Di Tommaso, 2025). Dans le cadre du projet CLIMATOR (Brisson and Levrault, 2010), des simulations de rendement du blé d’hiver dans des conditions climatiques futures ont été réalisées avec des modèles de culture sur différentes régions françaises. Dans les Hauts de France par exemple (Figure 1), bassin majeur de production de blé d’hiver, elles ont prédit une augmentation de production résultant de l’élévation en CO2, et ce malgré (i) un raccourcissement des cycles de croissance et une anticipation des stades phénologiques résultant du réchauffement climatique, (ii) une diminution du risque de gel, (iii) une augmentation des stress thermiques tardifs et (iv) un accroissement des déficits hydriques tardifs et précoces associés aux prédictions climatiques futures. Ces résultats issus du projet CLIMATOR ont depuis été confortés par d’autres études portant sur le calcul d’indicateurs écoclimatiques (Le Roux et al., 2024).
Exemple d’impacts simulés moyens du changement climatique sur la phénologie du blé d’hiver et sur les intensités ou fréquences de stress simulés dans les Hauts de France (d’après Brisson and Levrault, 2010). Les simulations ont été réalisées avec le modèle PANORAMIX (Arvalis), scénario SRES A1B correspondant au scénario médian SSP2-4.5 (concentrations de CO2 de 325, 415, 500 et 600 ppm en 1970, 2020, 2050 et 2100). Gel = risques de gel, DH = déficit hydrique en mm pour une réserve utile de 104 ou 193 mm, eT = nombre de jours avec des températures >25 °C entre l’épiaison et le stade laiteux, Echaud = nombre de jours échaudants pendant le remplissage.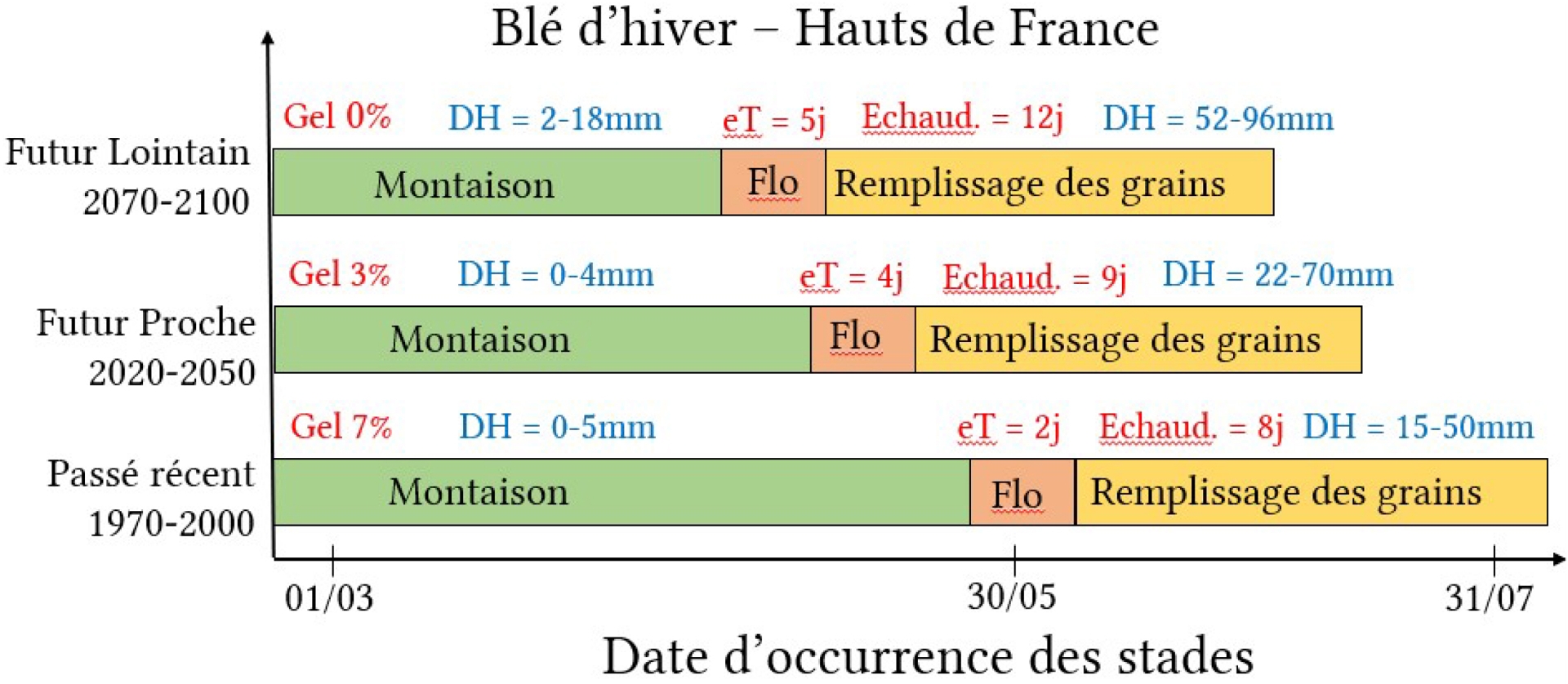
Cette étude illustre un effet bien connu du changement climatique, et en particulier de l’augmentation des températures, sur la phénologie avec un raccourcissement des cycles. Toutefois, cet effet peut être modulé par la date de semis et l’utilisation de variétés plus ou moins précoces (Minoli et al., 2022). Par ailleurs, cette étude souligne que la concomitance de différents évènements climatiques peut aboutir à des stress multiples qu’il est nécessaire d’estimer et de comprendre, en particulier avec des dispositifs expérimentaux ad hoc couplés à des approches de modélisation mécaniste. La question plus particulièrement abordée ici est de savoir, au vu des expérimentations menées, si l’effet fertilisant du CO2 compensera ou pas (i) le raccourcissement du cycle induit par l’augmentation tendancielle des températures et (ii) les effets des stress thermiques et hydriques croissants.
2. Méthodologies employées : des dispositifs expérimentaux au traitement de données
La question des impacts du changement climatique sur le rendement des cultures préoccupe la recherche internationale depuis plus de 40 ans. Elle a tenté d’y répondre en mettant en place divers dispositifs expérimentaux (Table 1) susceptibles d’apporter des références sur la réponse des plantes et peuplements à différents facteurs environnementaux clés, à savoir la température, la disponibilité en eau et le CO2, principalement, mais également l’ozone et les dépôts d’azote atmosphérique (Liang et al., 2023).
Dispositifs expérimentaux mobilisés pour analyser les impacts du changement climatique (incluant les effets du CO2) sur les écosystèmes cultivés ou naturels
| Type de dispositif | Herbarium, dispositifs longue durée | Sources naturelles CO2 | Chambres de culture, serres | Open-Top Chambers (OTC) | Free-Air CO2 Enrichment (FACE) |
| Facteurs contrôlés | Aucun | Aucun ou essais agronomiques | CO2 et autres facteurs (température, eau, azote) | CO2 et autres facteurs (température, eau, azote) | CO2 et autres facteurs (température, eau, azote) |
| Gammes CO2 (ppm) | 270–440 ppm | Ambiant +100 à +10 000 ppm, en moyenne 750 ppm | Jusqu’à 1000 ppm | Jusqu’à 1000 ppm | Ambiant +100 à 250 ppm soit jusqu’à 650 ppm |
| Interactions | Multiples Non contrôlées |
Multiples, non contrôlées | Binaires (ternaires) | Binaires à ternaires contrôlées avec facteurs non contrôlés | Multiples possibles |
| Gammes autres facteurs | Selon zone géographique | Selon zone géographique Témoins proches CO2 ambiant |
+1 à +4 °C, nutrition eau et azote extrême | +1 à +4 °C, nutrition eau et azote extrême | Gammes futures possibles en combinaisons, conditions naturelles multisites |
| Avantages | Tendances à long terme Multi-sites en conditions naturelles |
Impacts long terme sur populations naturelles Acclimatation long terme |
Essais factoriels contrôlés et constants, gammes futures possibles | Essais factoriels semi-contrôlés en conditions naturelles, gammes futures possibles, coût limité | Essais factoriels contrôlés avec répétitions, multisites en conditions naturelles |
| Inconvénients | Gammes passées Interactions multiples non quantifiées |
Fortes fluctuations CO2
Interactions multiples non quantifiées |
Peu de combinaisons analysées, conditions artificielles, effets de bord | Peu de combinaisons de facteurs, effets de bord et zone utile faible | Peu de combinaisons de facteurs, coût élevé, fluctuations de CO2 et gammes restreintes |
| Référence | Lang et al. (2024) | Saban et al. (2019) | Farkas et al. (2021); Varga et al. (2017) | Balouchi et al. (2009); Manderscheid and Weigel (2007) | Ainsworth and Long (2021); Allen et al. (2020) |
Ainsi, les conservatoires de plantes locales permettent de suivre sur de longues périodes en conditions naturelles les évolutions tendancielles de fitness (ou succès reproducteur), croissance ou rendement en réponse à l’augmentation du CO2 ; ces dispositifs multilocaux combinent également différents niveaux de stress, mais les niveaux de stress et de CO2 n’atteignent pas les niveaux prévus dans le futur, limitant ainsi leur intérêt en prédiction ; néanmoins ils présentent un grand intérêt pour confirmer des tendances sur le long terme. Des expérimentations en conditions semi-contrôlées (serre) ou contrôlées (chambre de culture) permettent de pallier ce problème en étendant et combinant des gammes de facteurs correspondant aux gammes futures ; néanmoins les combinaisons et modalités sont souvent restreintes, notamment à l’échelle de la plante, pour optimiser l’espace et les coûts. Si ces dispositifs permettent d’établir des normes de réaction utiles en modélisation, ils explorent en général un nombre faible de combinaisons de stress, et manquent souvent de réalisme pour des peuplements cultivés (Poorter, Fiorani et al., 2016). Trois autres types de dispositifs s’essaient à combiner gammes futures et expérimentations en conditions naturelles sur peuplements ou populations cultivées ou naturelles : sources naturelles de CO2, Open-Top Chambers (OTC) ou chambres légères à ciel ouvert permettant un enrichissement en CO2 en conditions extérieures, et Free-Air-CO2-Enrichment (FACE) ou surfaces expérimentales équipées d’un système continu d’injection du CO2 assurant l’atteinte d’une teneur cible. Les enrichissements à proximité d’une source naturelle de CO2 permettent de cibler les questions d’acclimatation au CO2 ambiant dans des écosystèmes étant soumis constamment à de fortes teneurs en CO2 (ambiant + 750 ppm en moyenne), et ce, avec des stress variables selon l’année et le site (Lang et al., 2024; Saban et al., 2019) ; cependant, les teneurs en CO2 sont très variables dans le temps et cette variabilité temporelle n’est typiquement pas prise en compte dans les impacts. Les OTC sont des chambres semi-ouvertes de tailles variables et applicables à des peuplements avec des teneurs en CO2 atteignant jusqu’à 1000 ppm, ordre de grandeurs comparables à ce qui pourrait être observé à la fin du siècle selon certains scénarios d’émissions de gaz à effet de serre ; cependant, ces systèmes semi-clos montrent des effets de bord limitant la surface utile homogène (De Alencar et al., 2024). Enfin, les FACE se développent depuis 30 ans et consistent en une injection de CO2 en système ouvert selon différents moyens et permettent de disposer d’une surface traitée importante en conditions naturelles (Ainsworth and Long, 2021) ; des modalités agronomiques peuvent y être étudiées (disponibilité en azote et eau essentiellement) ainsi que des effets conjoints d’augmentation (artificielle) de température de 1 à 4 °C en moyenne (T-FACE ; Fitzgerald et al., 2022). Les dispositifs FACE semblent les plus adaptés pour l’acquisition de références sur les peuplements cultivés. Cependant, ils ne permettent une élévation du CO2 que de 100 à 250 ppm par rapport au CO2 ambiant. De plus, les fluctuations rapides du CO2 observées dans ces dispositifs ouverts ne sont pas sans influence sur les réponses physiologiques mesurées, les minimisant de l’ordre de 35 % par rapport aux dispositifs sans fluctuation de CO2, tels que les OTC (Allen et al., 2020). Bien qu’aucun de ces types expérimentations ne soit sans limitation, on peut cependant considérer cette diversité de dispositifs comme un atout majeur pour valider ou non les changements d’échelle des impacts, qu’ils soient spatiaux, de l’organe à l’écosystème, ou temporels, de la minute au cycle, voire pluriannuels, pour les systèmes pérennes.
Du fait de la diversité de dispositifs, d’échelles spatio-temporelles et d’objectifs pour mieux comprendre les impacts du changement climatique sur les agroécosystèmes, les variables mesurées sont également de nature très différentes ; on peut distinguer (i) des variables de flux généralement estimées à large échelle géographique, comme les flux nets d’eau et de CO2 à diverses échelles temporelles (Walker et al., 2019) ; (ii) des variables de flux estimées à des échelles géographiques moyennes, du bassin de production à la région, comme les productivités nettes ou brutes, généralement biomasse totale ou rendement (Fitzgerald et al., 2022; Kimball, 2016; Rezaei et al., 2023; Tausz-Posch et al., 2020), et (iii) des variables de flux estimées de l’échelle foliaire à celle du m2 sur le court terme, comme des mesures de photosynthèse, respiration et transpiration (Dusenge et al., 2019) ; et enfin, (iv) des traits fonctionnels d’autre part, tels que la surface foliaire maximale, la conductance stomatique, les masses ou azote spécifiques (Poorter, Knopf et al., 2022). Cette diversité de variables rend délicates les comparaisons entre études.
Enfin, le traitement des données collectées dans les articles ou les méta-analyses introduit encore une autre source d’incertitudes pour dégager des tendances robustes de réponse aux combinaisons de facteurs/conditions liés au changement climatique. En effet, les données peuvent être exprimées (i) en valeur absolue, (ii) en valeur relative (Ainsworth and Long, 2021; Leakey et al., 2009; Toreti et al., 2020), mais avec une référence de CO2 ambiant qui varie selon les études, ou encore (iii) en valeurs normalisées (Liang et al., 2023; Norby, 2025; Walker et al., 2019). Ces différentes transformations font l’hypothèse d’une interpolation linéaire possible, ce qui est valide pour la réponse de croissance des plantes entre 300 et 900 ppm de CO2 (Idso, 1994; Kimball, 2016), mais difficile à soutenir pour la température au-delà des optimums thermiques. L’hétérogénéité d’expression des impacts bruite également les tendances générales dégagées, et suggère qu’une charte générale d’acquisition et de traitement des données, comme réalisée pour les traits en écologie (Joswig et al., 2021), permettrait une meilleure mutualisation et généralisation des résultats.
3. Fonctionnement des agroécosystèmes
La Figure 2 propose une représentation très simplifiée d’un peuplement végétal pour illustrer les effets propres de eT, eCO2 et des déficits hydriques sur le fonctionnement des agroécosystèmes ; à noter que, dans un cadre distinguant facteurs ressources et facteurs de contrôle, le CO2 agit principalement comme une ressource carbonée pour la croissance, tandis que la température régule la cinétique du développement et des processus, et l’eau occupe une position intermédiaire, étant à la fois un substrat de la photosynthèse et un déterminant du fonctionnement des stomates (Salisbury and Ross, 1992). On distingue les effets sur les variables de flux (photosynthèse et transpiration principalement) sous le contrôle de la conductance stomatique (gs) et sur les variables d’état du système : Leaf Area Index (LAI, m2 de feuilles par m2 de sol), biomasse et rendement en kg⋅m−2, état hydrique et minéral. Les stomates, au centre, régulent les flux de CO2 et vapeur d’eau par unité de surface foliaire entre végétal et atmosphère en modulant leur ouverture et leur densité selon les conditions climatiques, tandis que le LAI dimensionne ces mêmes flux à l’échelle du peuplement végétal.
Impacts de l’augmentation de température (eT), du CO2 (eCO2) et du déficit hydrique sur le fonctionnement des agroécosystèmes, d’après Debaeke et al. (2025), Gawinowski et al. (2025) et Liang et al. (2023). Les ellipses symbolisent des variables de flux, les rectangles des variables d’état, et les flèches des effets positifs (trait continu) ou négatifs (trait pointillé) reliant ces différents éléments. Abréviations : e = élévation ; P = précipitations ; T = température ; gs = conductance stomatique ; N = azote ; Rg = rayonnement global ; VPD = déficit de pression de vapeur d’eau ; LAI = Leaf Area Index ou indice foliaire (en m2 de feuilles par m2 de sol).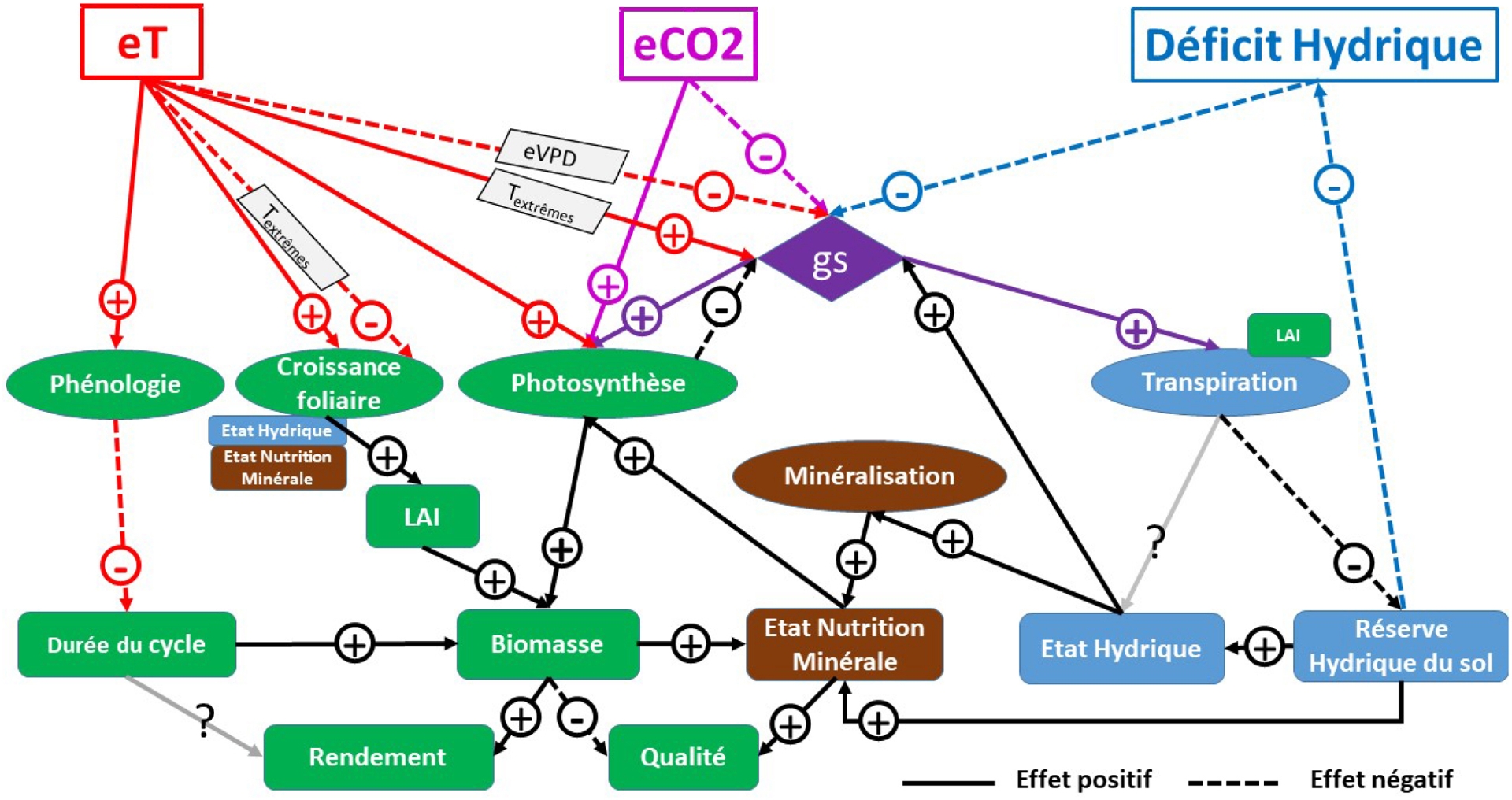
L’élévation de CO2 accroît la concentration interne en CO2 des cellules et augmente la photosynthèse, en particulier pour les plantes en C3 chez lesquelles la concentration en CO2 limite toujours le fonctionnement de la Rubisco ; cette augmentation du CO2 interne diminue la conductance stomatique (Long et al., 2004). L’accroissement de déficit hydrique diminue également la conductance stomatique, limitant la transpiration, ce qui limite le déstockage d’eau du sol par la plante, mais à l’inverse, limite les entrées d’eau dans le sol (Ahmad et al., 2018). Enfin l’élévation de température (eT) moyenne (en deçà de la température optimale Topt) impacte divers processus, et ce de manière parfois antagoniste sur la croissance. En effet, eT accélère le développement, raccourcit le cycle et donc la durée de l’acquisition de ressources, en particulier en CO2 (Girousse et al., 2021; Zheng et al., 2012) ; mais eT augmente l’initiation et la croissance foliaires (Gray and Brady, 2016), la surface foliaire spécifique (SLA en m2⋅kg−1, Dong et al., 2020), donc la taille des feuilles et le LAI, ainsi qu’observé à une échelle globale (Yang, Roderick et al., 2023). De plus, eT augmente la photosynthèse par unité de surface foliaire pour des températures inférieures à l’optimal, en particulier chez les plantes C3 tempérées. Par ailleurs, une augmentation des températures peut s’accompagner d’une augmentation du déficit de pression de vapeur saturante dans l’air, ce qui aboutit à une fermeture des stomates à partir d’un seuil donné (Bourbia and Brodribb, 2024). En cas de vagues de chaleur, où des températures extrêmes au-delà de l’optimum sont atteintes, les effets sur la conductance et la photosynthèse s’inversent : les stomates s’ouvrent, favorisant la transpiration et la diminution de température de surface des feuilles, mais inhibant le processus biochimique de photosynthèse (Farooq et al., 2011). En conséquence, ce schéma montre bien que les impacts résultant de ces facteurs et conditions liés au changement climatique sont très dépendants d’une part des réponses quantitatives individuelles aux facteurs ou conditions, et d’autre part, des priorités entre eux. Ainsi, Zandalinas and Mittler (2022) ont montré la prédominance des effets du déficit hydrique par rapport à ceux des températures extrêmes sur la conductance stomatique, mais variable selon les organes concernés (Singh et al., 2022).
Moyennant les remarques méthodologiques précédentes (Partie 3), la sensibilité moyenne à chacun des facteurs ou conditions, — à savoir l’eCO2 (par incrément de 100 ppm), l’eT (par °C < Topt) et le déficit hydrique — peut être estimée chez les plantes annuelles en C3 à partir des données d’articles de synthèse et de méta-analyses récentes (Ainsworth and Long, 2021; Fitzgerald et al., 2022; Jin et al., 2019; Kimball, 2016; Leakey et al., 2009; Norby, 2025; Rezaei et al., 2023; Saban et al., 2019; Toreti et al., 2020). L’absence de caractérisation homogène des déficits hydriques rend difficile tout calcul de sensibilité et généralisation entre espèces. Pour l’eCO2, la sensibilité sur différentes variables à une augmentation de 100 ppm de CO2 est remarquablement consensuelle, avec les ordres de grandeur suivants synthétisant l’ensemble des articles précédemment cités : de −8 à −9 % pour gs, +8 à +12 % pour la photosynthèse maximale (Asat), +13 % pour le LAI avec une diminution de surface spécifique entre −3,5 et −5 %, +8 à +10 % pour la biomasse et le rendement, une augmentation d’absorption d’azote de +4 à +12 %, ce qui conduit principalement à une diminution de −3 à −8 % des teneurs en protéines des organes récoltés pour une augmentation de 100 ppm. Par contre, l’évapotranspiration du peuplement diminue de −3 à −5 %, ce qui est d û à des augmentations de LAI et une diminution de conductance stomatique ; l’efficience de l’utilisation de l’eau augmente fortement, de l’ordre de 28 % en moyenne. Il est notable que les augmentations de Asat et biomasse des ligneux en réponse à l’eCO2 sont 2 à 3 fois plus importantes que celles des annuelles. Enfin, quelques méta-analyses (Ainsworth and Long, 2021; Kimball, 2016) permettent de classer la sensibilité des espèces au CO2 selon des pseudo types fonctionnels ; ainsi elles montrent que les C3 à tubercules sont plus sensibles que les C3 légumineuses, elles-mêmes plus sensibles que les autres C3 annuelles. Les C3 sont plus sensibles que les C4, qui répondent peu ou pas au eCO2. Enfin, les pérennes, qu’elles soient herbacées ou ligneuses, sont plus sensibles que les annuelles au eCO2.
Les données de sensibilité thermique sont moins nombreuses à cette même échelle, même si les normes de réaction de processus fins à la température sont nombreuses. Ainsi, la gs diminue en moyenne de −1,5 % par °C (<Topt), Asat augmente +4,8 % tout comme la biomasse et le rendement (+5 %), ce qui est cohérent avec l’augmentation de LAI avec la température généralement observée (Dong et al., 2020; Fitzgerald et al., 2022; Gray and Brady, 2016; Rasul et al., 2020; Yang, Roderick et al., 2023). A noter que la réponse à eCO2 et eT va souvent dans le même sens, contrairement à la réponse au déficit hydrique.
Quelques méta-analyses récentes ont également synthétisés les effets interactifs entre deux variables (eCO2, eT et/ou déficit hydrique), sur la conductance stomatique gs (Liang et al., 2023, sur un large échantillon de biomes) et sur le rendement (Ainsworth and Long, 2021). Si Liang et al. (2023) concluent à un effet globalement additif des facteurs (ou conditions) sur gs, les résultats sont plus variés sur le rendement (Ainsworth and Long, 2021) ; en particulier chez les C4, ces auteurs montrent clairement une absence d’effet de eCO2 sur le rendement de peuplements bien alimentés en eau alors qu’ils rapportent un rendement augmenté de 10 % pour +100 ppm de CO2 en cas de déficit hydrique. Ils montrent également des effets d’interaction moindres, mais notables avec eT et le déficit hydrique chez les C3.
En conclusion, étant donné la grande diversité des espèces, des dispositifs, des mesures réalisées avec des combinaisons de facteurs et conditions variables et caractéristiques des conditions climatiques futures, il est difficile de répondre de manière certaine à la question initiale « l’eCO2 compensera-t-il l’augmentation des stress hydriques et thermiques futurs ? », question plus spécifiquement traitée dans la Partie 5.
4. Synthèse des études expérimentales sur le blé
Dans cette section, nous nous intéressons plus en détail aux effets des interactions entre CO2, températures et déficit hydrique sur le blé, en nous appuyant sur la revue de (Gawinowski et al., 2025). Cette revue réalise une synthèse des expérimentations combinant une élévation de CO2 (eCO2) avec un déficit hydrique (WD, pour « water deficit ») et/ou une augmentation de température. Cette augmentation de température peut s’exprimer comme une hausse des températures moyennes (HT, pour « high temperatures ») ou comme un événement ponctuel tel qu’une vague de chaleur (HW, pour « heatwave »). Les expérimentations considérées sont réalisées dans différents types de dispositifs : chambres de culture, chambres open-top et FACE. Deux types de traits d’intérêts sont étudiés dans cette revue : les traits liés à la productivité, comme le rendement, la biomasse ou la photosynthèse, et les traits liés à la consommation en eau, comme la conductance stomatique, la transpiration ou l’efficience d’utilisation de l’eau. Les principales informations sur ces expérimentations sont regroupées dans le Tableau 2.
Méta-données des expérimentations recensées dans (Gawinowski et al., 2025) combinant augmentation de CO2 (eCO2), déficit hydrique (WD), hautes températures (HT) et vagues de chaleur (HW)
| Conditions climatiques | Nombre d’études | Nombre d’observations sur la productivité | Nombre d’observations la consommation en eau | Nombre de variétés de blé |
|---|---|---|---|---|
| eCO2 × WD | 29 | 79 | 57 | 41 |
| eCO2 × HT | 23 | 49 | 6 | 23 |
| eCO2 × WD | 9 | 40 | 31 | 28 |
| eCO2 × WD × HT | 5 | 21 | 3 | 7 |
| eCO2 × WD × HW | 3 | 8 | 8 | 8 |
Dans cette revue, des indices de plasticité relative sont calculés pour évaluer les interactions entre le CO2 et d’autres facteurs climatiques, mais aussi les compensations entre ces facteurs en regardant si les effets de stress (température et eau) peuvent être compensés par une élévation de CO2 (Figure 3). Les indices de plasticité sur les interactions répondent à la question « Est-ce que l’effet du CO2 sur un trait d’intérêt varie avec le niveau de stress (température et/ou eau) ? » tandis que les indices de plasticité sur la compensation répondent à la question « Quelle est la variation du trait d’intérêt entre (i) des conditions ambiantes en CO2 et non stressantes, et (ii) des conditions futures avec augmentation du CO2 et des stress ? »
Schéma conceptuel de la définition d’indices de plasticité pour la compensation et l’interaction entre le CO2, le déficit hydrique et les fortes températures (adapté de Gawinowski et al., 2025). Ces indices sont des variations relatives calculées pour deux types de traits liés à la productivité (PRT) ou la consommation en eau (WRT). Les ΔCO2 représentent la variation de ces traits entre des concentrations de CO2 ambiantes et élevées, dans des conditions non stressantes (NS) ou stressantes (S, hautes températures et/ou déficit hydrique). Les indices d’interaction (en violet) comparent ces ΔCO2 en conditions NS ou S. Les indices de compensation (en orange) indiquent l’évolution directe des traits entre des conditions ambiantes actuelles et des conditions « futures » (eCO2 et stress).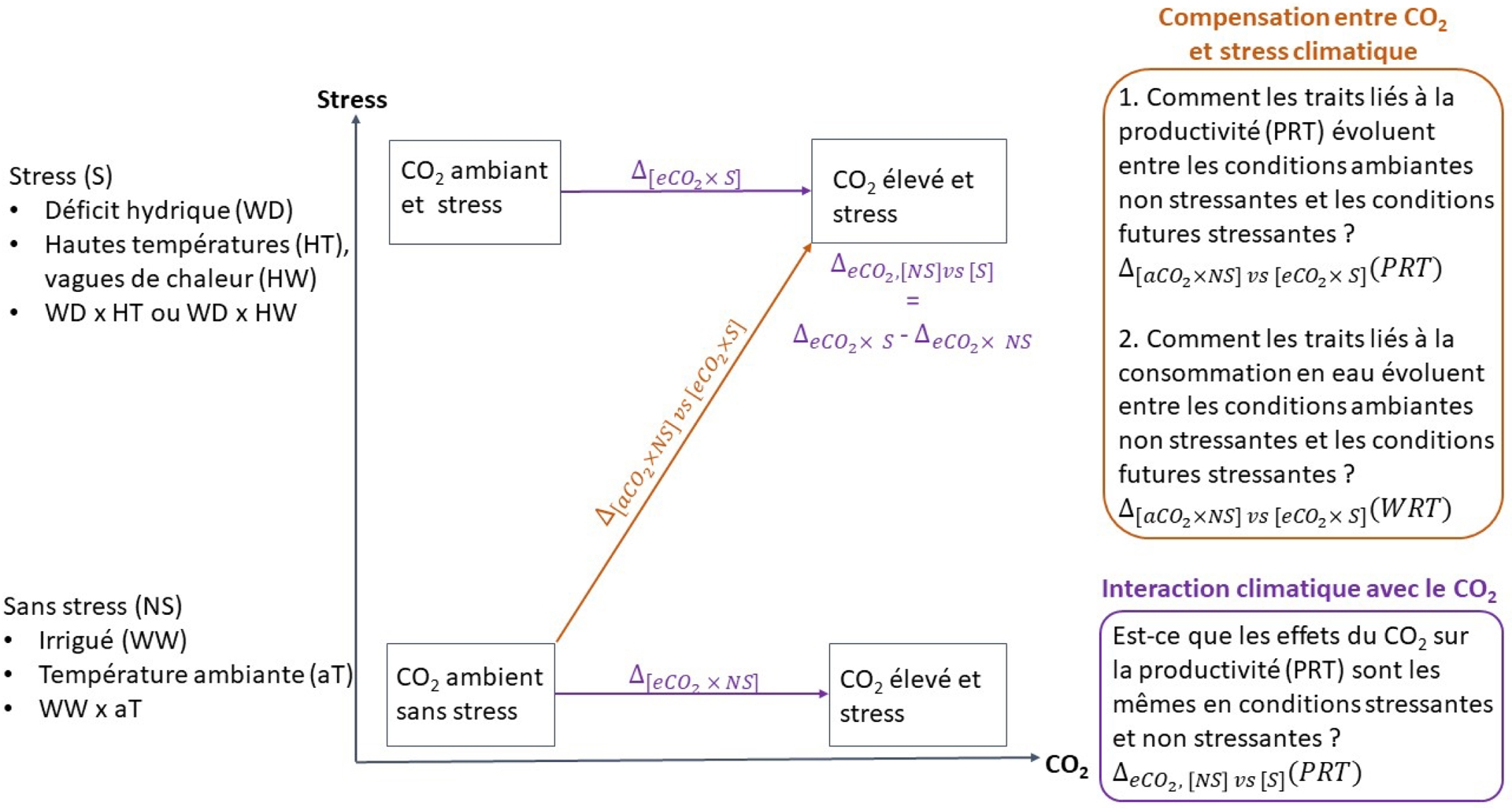
Dans cette revue, les indices de plasticité relative sont représentés sous forme de normes de réaction (par exemple, Figure 4 pour la compensation entre CO2 et déficit hydrique). Cette représentation permet de visualiser directement l’ensemble des réponses pour chaque génotype et condition expérimentale, en distinguant les différents dispositifs, les types de traits, l’augmentation de CO2 et l’intensité du stress. Pour une modalité expérimentale donnée (une variété, une année, une modalité d’irrigation ou de température, pour une étude donnée), une norme de réaction est représentée entre les deux concentrations de CO2 expérimentées en abscisse, avec une variation relative à 0 % pour la concentration ambiante et la valeur de l’indice de plasticité pour la concentration élevée. Par exemple, un indice de compensation à +25 % sur la Figure 4 indique une augmentation de 25 % de la productivité dans les conditions « futures » eCO2 et déficit hydrique par rapport aux conditions actuelles aCO2 et absence de stress hydrique. Ainsi, la Figure 4 présentant la compensation entre CO2 et déficit hydrique sur les traits de productivité montre que les expérimentations considérées sont déséquilibrées en nombre et en intensité entre les différents dispositifs, avec des réponses relatives très variables. Une diminution générale de la productivité est observée dans 55 cas sur 79 (normes de réaction négatives), avec une diminution moyenne de 9 %, indiquant que l’augmentation de CO2 ne permet pas de compenser les effets négatifs du déficit hydrique sur la productivité pour les conditions expérimentales étudiées. De plus, dans les dispositifs OTC et FACE, bien que les intensités de déficit hydrique soient modérées, les réponses de la productivité sont majoritairement négatives.
Normes de réaction des variations relatives (%) des variables de productivité (biomasse végétative, rendement) entre des conditions ambiantes/humides (aCO2, WW) et des conditions futures/sèches pour le CO2 et le déficit hydrique (eCO2, WD). Les résultats sont présentés par type de dispositif (chambre, OTC, FACE). Chaque norme de réaction correspond à une modalité expérimentale avec en abscisse les concentrations de CO2 expérimentées. Les couleurs représentent l’intensité du déficit hydrique imposé dans les expérimentations (ratio des apports d’eau entre traitements, %) et le type de ligne représente le stade d’exposition au déficit hydrique. Les lignes noires pointillées horizontales indiquent 0 %. Figure adaptée de Gawinowski et al. (2025).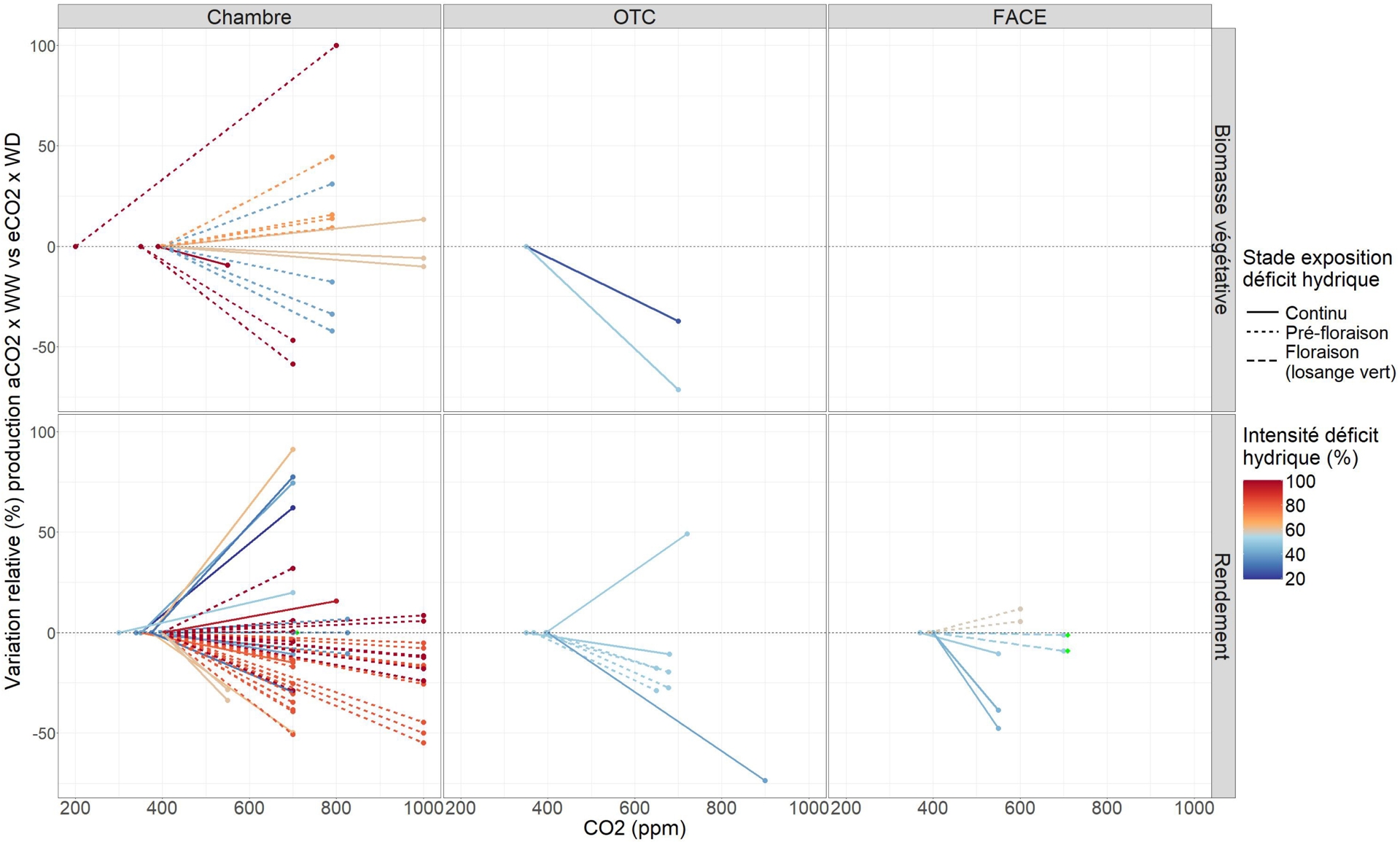
Les normes de réaction pour les deux types de traits d’intérêt, productivité et consommation en eau, sont présentées en détail pour les différentes conditions climatiques (eCO2 × WD, eCO2 × HT, eCO2 × HW, eCO2 × WD × HT et eCO2 × WD × HW) et sont dans la revue de (ibid.). Les informations générales sont représentées sur la Figure 5. Concernant la compensation sur la productivité (Figure 5A), dans l’ensemble des combinaisons climatiques expérimentées, une tendance générale de diminution de la productivité se confirme. Bien que les conditions de eCO2 et de hautes températures montrent une variation quasi nulle (+0,03 %) par rapport aux conditions ambiantes sans stress, l’effet le plus délétère est observé dans les conditions de eCO2 combiné avec du déficit hydrique et de hautes températures (−25 %). Dans ces conditions de eCO2 et de vagues de chaleurs, il semble que le stade phénologique durant lequel est subi le stress est plus impactant que la température ou la durée, avec un stress lors de la préfloraison plus délétère, mais avec un possible effet confondant lié à la tolérance à la chaleur des différentes variétés utilisées (voir Gawinowski et al., 2025). Dans les conditions combinant eCO2, déficit hydrique et températures élevées, malgré un faible nombre de données (21 pour HT et 8 pour HW, Figure 5A), il semble que la tolérance variétale à la sécheresse soit plus bénéfique que la tolérance à la température, en accord avec les conclusions de Zandalinas and Mittler (2022) sur la dominance du stress hydrique sur le stress thermique.
Variations relatives (%) des indices pour la compensation sur la productivité, et sur la compensation sur la consommation en eau, ainsi que variations relatives des indices d’interaction entre l’augmentation de CO2 et le déficit hydrique (WD) et/ou la température, via des hautes températures (HT) ou des vagues de chaleur (HW). Les barres représentent les valeurs moyennes sur l’ensemble des expérimentations, et les barres d’erreurs représentent les erreurs standards. Les boîtes représentent le nombre d’observations pour chaque condition. La base de données utilisée est celle de Gawinowski et al. (2025).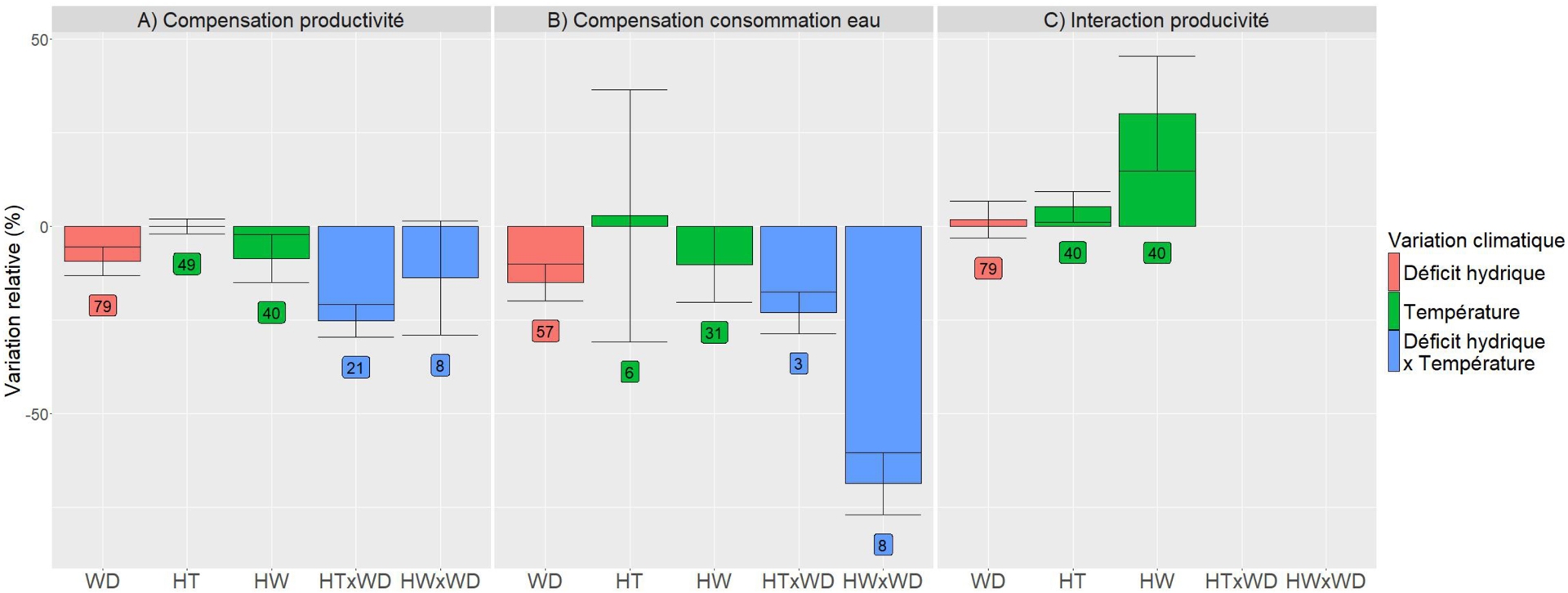
Une tendance générale de diminution se dessine également pour la consommation en eau (Figure 5B) : difficile à estimer dans les conditions eCO2 et hautes températures (peu d’observations, et forte variabilité) avec un effet moyen de −10 % pour les conditions eCO2 et vagues de chaleur, −25 % pour les conditions eCO2 avec déficit hydrique avec/sans hautes températures, et une diminution de −70 % dans les conditions de eCO2, déficit hydrique et vague de chaleur. On constate donc que les effets du CO2 et du déficit hydrique diminuent la consommation en eau, notamment dans des conditions de hautes températures ; par ailleurs, cette diminution est plus importante pour les vagues de chaleur que pour les hautes températures.
L’impact du CO2 sur la productivité a également été analysé en fonction de l’interaction avec le déficit hydrique et les températures (Figure 5C). Selon une hypothèse majoritaire dans la littérature, l’effet fertilisant du CO2 serait plus important en conditions sèches qu’en conditions humides ; or, sur l’ensemble des études considérées, les résultats sont très variables et l’hypothèse n’est vérifiée que dans la moitié des cas. Une interaction positive entre le CO2 et la température a également été constatée pour 55 % des cas pour les hautes températures, et 69 % des cas pour les vagues de chaleur. Cette interaction positive est cohérente avec les hypothèses de Long (1991) et (Qaderi and Reid, 2009) d’une augmentation de l’optimum de température de la photosynthèse avec l’augmentation de CO2.
Malgré les tendances générales qui peuvent être tirées de cette revue, pour de nombreuses conditions expérimentales, ces résultats moyens sont accompagnés d’une variabilité très importante et sont souvent estimés sur un faible nombre d’observations. Par ailleurs, les expérimentations comparées sont réalisées dans des conditions très différentes : sites, climats, sols, culture en pot ou en plein champ, conditions contrôlées ou conditions variables, échelles et fréquences de mesure des variables d’intérêt, processus ciblés, timing/intensité/durée des stress …
En particulier, la diversité génétique mobilisée est conséquente avec un grand nombre de variétés ; ces variétés sont souvent choisies en adéquation avec les contextes locaux des différentes expérimentations avec une distribution géographique mondiale (Europe, Chine, États-Unis, Australie), notamment pour leur tolérance aux hautes températures ou à la sécheresse.
Par ailleurs, il est difficile de caractériser les différents stress de façon standardisée entre ces expérimentations, ce qui pourrait être amélioré avec l’emploi d’indicateurs écoclimatiques (Le Roux et al., 2024). D’autre part, il a été souligné dans Zhu et al. (2023) que les expérimentations combinant CO2 et hautes températures étaient réalisées dans des conditions climatiques souvent disjointes des projections climatiques, en particulier à long voire très long terme et pour des scénarios d’émissions élevées. Il est donc nécessaire de produire un plus grand volume de données en accès libre dans des conditions climatiques pertinentes par rapport aux projections climatiques futures, avec une caractérisation standardisée des différents stress.
Les parties 4 et 5 montrent les effets convergents du eCO2, des fortes températures et déficits hydriques, avec des interactions entre ces facteurs et une absence de compensation systématique des stress hydriques et thermiques par l’effet fertilisant du CO2. La généralisation de ces effets à l’ensemble des biomes reste toutefois limitée par une forte variabilité entre études, liée à la diversité des types fonctionnels (types métaboliques C3 ou C4 ; annuelles et pérennes ; légumineuses et non légumineuses ; rendement en biomasse, en grains ou en tubercules, par ex.), et aux différentes conditions pédoclimatiques entre localisations géographiques et dates d’études (Norby, 2025). Les réponses quantitatives dépendent également (i) des dispositifs expérimentaux et des gammes de variations qu’ils permettent (ainsi, les réponses obtenues sont de moins en moins fortes selon les dispositifs) ; et (ii) de la durée des expérimentations : une acclimatation à long terme est constatée, faible sous eCO2, mais plus marquée sous réchauffement (Cai et al., 2016; Saban et al., 2019). Malgré cela, la cohérence des sens de variation de leurs ordres de grandeur entre les différents dispositifs OTC, FACE et enrichissement naturel renforce la robustesse des résultats présentés, en l’absence de stress biotiques, plus particulièrement abordés dans la partie suivante.
Très peu d’études historiques considèrent et quantifient les impacts du changement climatique sur les composantes biotiques associées aux cultures, annuelles ou pérennes. Ces impacts pourraient cependant modifier la productivité à court, moyen ou long terme, en particulier dans des systèmes à faibles intrants, qui dépendent de fait davantage des régulations biologiques pour assurer leur résilience face au changement climatique. Ces aspects sont très brièvement abordés par quelques exemples dans une dernière partie de cette revue, sans rechercher l’exhaustivité, mais comme une ouverture à des résultats qui pourraient altérer les tendances précédentes.
5. Changement climatique et stress biotiques dans les agroécosystèmes
Les composantes biologiques de l’agroécosystème les plus fréquemment étudiées en réponse au changement climatique actuel sont (i) les bioagresseurs (Debaeke et al., 2025), du fait de leur nuisibilité (pertes de rendement allant de 25–35 % à 50–80 % avec et sans traitement ; Oerke, 2006) et de leur potentiel invasif, de dispersion et d’adaptation aux conditions environnementales ; et (ii) le microbiote du sol (Yang, Li et al., 2021), très réactif aux modifications du sol et de l’environnement, qui est une composante majeure de la santé des sols et cultures. Les bioagresseurs des cultures sont majoritairement des microorganismes pathogènes (champignons, bactéries et virus), des insectes (arthropodes principalement) et des plantes (adventices), principalement ectothermes, souvent de petite taille, à cycle court et à taux de multiplication élevé. Les facteurs climatiques, en particulier la température et l’humidité de l’air, qui varient en fonction du changement climatique, vont affecter directement leur cycle de vie, pilotant la faisabilité de l’infection (combinaison propre de température et durée d’humectation ; Magarey et al., 2005) et leur phénologie (Delmas et al., 2024). Le nombre de cycles est un déterminant majeur de l’épidémie, mais leur dispersion sera également modifiée, en particulier à longue distance, via d’autres facteurs du changement climatique non abordés (vents, tornades, cyclones, etc. ; Seidl et al., 2017). Les larges variations inter- et intra-spécifiques de normes de réactions observées (Magarey et al., 2005) conduisent à des variations de profils de bioagresseurs avec le changement climatique, à travers un filtrage saisonnier ou géographique des espèces ou génotypes les plus adaptés localement (Suffert et al., 2018). Une fois dans les tissus, la température reste le facteur déterminant majoritaire de l’épidémie, même si le eCO2, conduisant à des surfaces sensibles plus grandes et des tissus plus appétants, favorise la croissance des organismes biotrophes (Leisner et al., 2023). Enfin, le climat joue un rôle prépondérant dans la survie des propagules infectieuses entre les cycles de culture ; en particulier la diminution des gels hivernaux avec le changement climatique accroît le stock d’inoculum pour des infections de printemps plus précoces et intenses (ex. Xylella fastidiosa, Godefroid et al., 2022). L’ensemble de ces effets directs du climat sur les bioagresseurs des cultures aboutit à une augmentation des zones géographiques où apparaissent de nouveaux bioagresseurs (Bebber, 2015), dont 40 % sont attribués au changement climatique (Corredor-Moreno and Saunders, 2020), et à un déplacement géographique moyen vers les pôles de 2,7 km/an depuis 1960 (Bebber et al., 2013), mais qui est assez variable selon les groupes taxonomiques. Enfin, ces organismes à cycle court ont un fort potentiel d’adaptation aux conditions climatiques, en particulier à l’échelle populationnelle ; ainsi, la température optimale des populations de Zymoseptoria. tritici, agent causal de la septoriose du blé, varie entre automne et printemps (Boixel et al., 2022), tout comme les fréquences d’isolats adaptés à un milieu chaud ou froid augmentent sous des régimes thermiques respectivement chauds et froids (Boixel, 2020). Ces adaptations populationnelles peuvent conduire à des extensions géographiques rapides en conditions suboptimales.
Ces effets directs sont largement accompagnés d’effets indirects à travers les habitats plus ou moins favorables que génère le changement climatique. En effet, la réponse des organismes à la température étant très variable, une augmentation des températures peut générer des différences de phénologies telles que les phases sensibles des plantes ne correspondent plus à l’arrivée des bioagresseurs, comme dans le cas de la moniliose de l’abricotier (Tresson et al., 2020). Ces disjonctions de niches écologiques peuvent avoir des répercussions en cascade sur le fonctionnement des écosystèmes (ex. arbres, arthropodes et oiseaux), en termes de ressources trophiques (Renner and Zohner, 2018). Si une augmentation des températures permet une faisabilité accrue vers le nord des cultures, elle accroît également la sensibilité des plantes à certains parasites (Chaloner et al., 2021; Desaint et al., 2021), tandis qu’une augmentation de CO2 augmente la disponibilité en organes sensibles, mais également leur teneur en composés de défense (Hamann et al., 2021). Ces interactions multiples et parfois contradictoires sont particulièrement visibles à travers les effets du changement climatique sur la fermeture stomatique (Lin et al., 2022), d’une part défavorable à l’entrée des pathogènes ou ravageurs dans les tissus d’une part, et d’autre part défavorable au développement d’une immunité collective en limitant les émissions de composés organiques volatils (COVs) « prévenant » les tissus ou plantes voisines. Ainsi, Hamann et al. (2021) illustrent bien par méta-analyse ces effets directs et indirects dans le cas des effets propres et en interactions des augmentations de CO2 et des températures, et de la sécheresse sur des populations d’herbivores : la baisse de qualité nutritionnelle des tissus accroissent à court terme la consommation et donc l’impact des herbivores sur les cultures, mais limitent leur fécondité et donc leur abondance à plus long terme. Ces interactions multiples en cascade, assorties d’une forte variabilité, rendent la prédiction de la nuisibilité de complexes de bioagresseurs très incertaine (Fitzgerald et al., 2022).
Les interactions entre peuplement végétal et environnement à travers le microbiote du sol sont de plus en plus étudiées, et en particulier le rôle de ce dernier dans la santé des plantes (Pélissier et al., 2021), si bien que Singh et al. (2022) suggèrent de l’intégrer comme axe supplémentaire du triangle épidémique plante-pathogène-environnement classique chez les épidémiologistes. Les impacts des interactions entre facteurs/conditions du changement climatique sur le microbiote le sont moins. Citons simplement la récente méta-analyse de Yang, Li et al. (2021) qui synthétise sur ces aspects les résultats de 237 articles scientifiques menés en multisites. Ils constatent un effet majoritairement négatif du changement global sur l’abondance et la diversité des communautés microbiennes du sol, bactériennes comme fongiques, malgré un effet propre positif des augmentations de CO2 et des températures. Les interactions entre 2 à 3 facteurs aggravent largement les effets propres négatifs, mais montrent également, dans le cas de la combinaison eCO2 et eT, un impact négatif significatif sur la biodiversité bactérienne. Ces impacts jouent probablement sur la santé et la productivité des écosystèmes naturels ou cultivés à moyen et long termes, ce qui demande des approfondissements futurs. Ce paragraphe montre que mieux connaître la résilience des agroécosystèmes au changement climatique nécessite non seulement de mieux appréhender les impacts de facteurs et conditions en interactions sur le fonctionnement des peuplements végétaux, mais également, plus largement, sur les composantes biologiques associées qui peuvent fortement altérer leurs productivité, qualité et durabilité.
6. Perspectives pour la modélisation des cultures
L’étude de la plasticité des plantes face aux variations environnementales par des approches expérimentales est limitée par la combinaison de modalités de stress et de pratiques culturales que l’on peut tester in situ. Cependant, ces approches peuvent être élargies in silico par l’utilisation de modèles de culture pour évaluer et explorer les impacts du changement climatique sur les cultures et prédire les niveaux de production dans des conditions climatiques futures (Asseng et al., 2019; Chenu et al., 2017; Collins and Chenu, 2021). Cependant, des interrogations ont été soulevées quant à la validité des résultats de simulation de ces modèles en contexte de changement climatique, telles que la surestimation des effets du eCO2 (Asseng et al., 2019) ou la sous-estimation des stress thermiques et hydriques combinés (Chenu et al., 2017). Des ensembles de modèles de culture sont mobilisés dans le cadre de l’initiative internationale d’intercomparaison AgMIP, la médiane d’un ensemble de modèles étant un bon prédicteur de la réponse des cultures au changement climatique (Asseng et al., 2019), bien que le nombre de conditions climatiques testées reste faible. Toutefois, cette médiane cache une variabilité très importante entre les différents modèles, qui peuvent surestimer ou sous-estimer des variables clés. Des études récentes concluent ainsi que les modèles de cultures ont tendance à sous-estimer les effets négatifs du changement climatique sur les rendements des cultures (Nóia Júnior et al., 2025), en particulier les effets des sécheresses, avec une sous-estimation systématique de l’évapotranspiration (Webber et al., 2025). Une validation rigoureuse et systématique de ces modèles sur des données expérimentales combinant eCO2, températures et déficit hydrique est donc essentielle, mais pose différentes difficultés liées à l’acquisition et à l’accès aux données expérimentales, mais aussi aux méthodologies de calibration/validation des modèles sur des données hétérogènes. La prise en compte des processus à l’échelle de l’organe pourrait constituer une source d’amélioration des modèles de culture dans cette perspective, comme ce qui a été fait pour APSIM (Brown et al., 2014) ou le modèle structure-fonction CNW-Wheat (Acker, 2025).
7. Conclusion générale
Le changement climatique modifie simultanément plusieurs facteurs et conditions clés du fonctionnement des agroécosystèmes, au premier rang desquels l’augmentation de la concentration atmosphérique en CO2, l’élévation des températures et l’intensification des déficits hydriques. Cette synthèse montre qu’au-delà des effets propres de ces facteurs, les effets de leurs interactions restent difficilement généralisables en raison de la diversité des dispositifs expérimentaux, des échelles d’étude et des contextes biophysiques considérés. À l’échelle des peuplements cultivés, l’effet fertilisant du CO2 améliore globalement l’efficience d’utilisation de l’eau et certains processus de croissance, mais ne permet pas, de manière systématique, de compenser les effets négatifs des stress hydriques et thermiques, en particulier lorsqu’ils sont combinés et/ou surviennent à des stades sensibles du cycle de culture.
La synthèse des études sur le blé met en évidence une forte variabilité des réponses, dépendante à la fois des conditions climatiques imposées, des dispositifs expérimentaux et de la diversité génétique considérée. Elle souligne la dominance fréquente de l’impact du déficit hydrique sur les hautes températures dans les réponses de consommation en eau, ainsi que l’absence de compensation généralisée par le CO2 sur la productivité dans les conditions climatiques futures expérimentées. Au-delà du fonctionnement des plantes cultivées, le changement climatique affecte également les composantes biologiques associées aux agroécosystèmes, notamment les bioagresseurs et le microbiote du sol, introduisant des rétroactions complexes susceptibles d’amplifier ou de moduler les impacts directs sur les rendements et la durabilité des systèmes agricoles. Dans ce contexte, le développement de dispositifs expérimentaux combinant des gradients climatiques pertinents, une caractérisation des stress et un accès élargi aux données apparaît essentiel pour améliorer la robustesse des synthèses, mais aussi l’évaluation des modèles de culture. L’intégration explicite des interactions entre facteurs climatiques, des processus biologiques associés et de la variabilité génétique constitue un levier majeur pour mieux anticiper les trajectoires de productivité des agroécosystèmes dans un climat en changement.
Remerciements
Nous remercions le projet FSOV REGARD (« RechErche d’analoGues climAtiques pour sélectionneR Demain »), coordonné par Arvalis, qui a financé ce travail de 2022 à 2025.
Déclaration d’intérêts
Les auteurs ne travaillent pas, ne conseillent pas, ne possèdent pas de parts, ne reçoivent pas de fonds d’une organisation qui pourrait tirer profit de cet article, et n’ont déclaré aucune autre affiliation que leurs organismes de recherche.