

1 Introduction
Recently one could observe an upsurge of contributions concerning impacts of Marine Protected Areas (MPAs). This growing interest for MPAs can be explained first by the various benefits expected from the creation of MPAs. These benefits can be broadly classified under three kinds: ecosystem preservation, fisheries management and development of the nonextractive recreational activities (Boncoeur et al. [2]). Secondly, MPAs are often presented as a new tool to control over-exploitation of the marine resource, which is a serious worldwide problem. Third, another motivation is that many MPAs have already been practiced all over the world. Lauck et al. [10] assert that MPAs can be envisaged as a kind of insurance against scientific uncertainty or stocks assessments or effectiveness of regulation errors.
Similar to Arnason [1], our paper is concerned with the study of theoretical conditions under which MPAs may be also economically beneficial. In biological perspective, MPAs generally increase abundance and average size of exploited species within their boundaries. There is an evidence that benefits may be exported to surrounding regions in some cases. We have restricted our attention to impacts on the fishing sector under the assumption that fishery is optimally managed by fishery managers (whether individuals or committees). In such a perfect world, we show that creation of a well designed MPA can improve biological situations within MPA boundaries, as well as economic benefits from the fishing activity outside its boundaries.
Following Hannesson [7], we use here a model where a given fishing ground is split into two sub-areas. One of them is set aside as a MPA. It is assumed that fish stock within the boundaries of MPA and stock fish into the surrounding area are linked by migration patterns of the resource. This framework is a variant of the spatial model developed by Sanchirico and Wilen [13]. This setting appears to be powerful and leaves open many possibilities with regard to what kind of migration may take place between stocks. It might be used for modeling a wide-ranging scale of problems. For instance, discussing the existence and the stability of an optimal harvesting policy (Dubey et al. [6]), or determining the optimal size of a MPA in the stochastic case (Conrad [5]).
Obviously, existence of economic benefits from the fishing activity outside MPA boundaries is contingent on the nature of biological linkages between areas. It will be assumed here that biological linkages are mainly dependent on adequate design of the MPA. By design we mean essentially here location and size. These are characteristics of major importance to appraise a migration coefficient that gives us some information on the fish mobility (cf. Houde [9]). In the present paper, it is then assumed that the knowledge of the migration coefficient allows one to identify and then choose the locations that have the highest potential for the implementation of a MPA. Therefore, the migration coefficient can be considered as a decision variable of fishery managers.
More precisely, the basic harvesting model, as it can be found, for instance, in Clark [4], is used as a benchmark. Though our study appears also as an optimal harvesting problem, we work, as Clark did, in the calculus of variation framework. Indeed the interior solutions of the optimal control problem, the only ones we are interested in, are straightforwardly obtained in this setting.
A MPA consideration is added to the basic model, leading to a model of two patchy populations that allows us to explore the possible effects of MPA. The biological and economics impacts, both within and beyond the borders of the MPA, are highlighted.
Optimal steady states, expressed in terms of fishing effort and stock density, are determined in both the basic and the patchy models. For the latter one, the optimal migration coefficient, i.e., the value where potential effect of MPA implementation should be highest, is obtained. Optimal benefits, with or without MPA, are compared. Conditions under which a MPA creation will theoretically enhance both biological (i.e. the stock density) and economic (i.e., the present value of the exploitation) situations are obtained. Unfortunately, it is likely uneasy to reach in practice the optimal value of the migration coefficient. Some simulations are therefore used to explore the sensitivity of our results with respect to the migration coefficient. It is shown numerically that the domain of the parameter for which the presence of a MPA is theoretically beneficial is reasonably large.
The paper is organized as follows. In Section 2 the basic fishery model is presented. In Section 3 the MPA component is added to the model. The implications for economics benefits and for the biomass are then explored. In Section 4 a set of results of simulations is shown. The last section summarizes the major conclusions and suggest some additional lines of research, for which is should be taken into account additional arguments in favor of MPA implementations.
2 The basic fishery model
Consider a fish stock distributed over a given area that we represent by its density X (defined as the ratio of the stock over the carrying capacity of the area). In accordance with classical modeling, the growth of the biomass density is given by:
| (1) |
The standard economic theory claims that fishery managers maximise the profits from harvesting, which amounts to deal with the following optimization problem
| (2) |
| (3) |
According to (2), for all trajectories s.t.
| (4) |
| (5) |
2.1 Optimality conditions
For the study of the solutions of (5), we introduce a function
| (6) |
If there exists a solution
2.2 Optimal fishing effort and fishery profit
When an optimal stationary solution
| (7) |
If
3 Marine protected areas and optimality
Let us consider that we have an optimally managed fishery and examine the impacts of the introduction of a MPA in such a situation. We claim that under an efficient fisheries management system, a MPA, properly defined and implemented, may enhance both economic benefits and fish stocks.
The model used here is structurally very similar to Hannesson's model [7]. It deals with sub-populations distributed in two patches interacting through migration. This is a variant of the spatial model developed by Sanchirico and Wilen [12].
It is assumed that the migration depends only on the biomass densities in each area (i.e., the ratio of stock over carrying capacity). The simplest migration model is based on diffusion, which depends merely upon the difference between the respective densities of patches. Therefore migration occurs if a disparity arises between the respective biomass densities inside and outside the MPA.
Following Conrad [5], Hannesson [7], it is presumed that the carrying capacity is increasing with the patch size. Nevertheless, we assume here that the carrying capacity of the MPA is always very small compared to the overall carrying capacity, which is equivalent to claim that the carrying capacity of the unprotected area is (almost) not modified by the existence of a MPA. Consequently, we consider here that the value P of the carrying capacity in the harvesting area is not modified by the creation of a MPA.
Suppose also that this spatial consideration allows us to distinguish the population behavior between two dynamics, as follows. The growth of the sub-population density into the MPA is governed by the dynamics:
| (8) |
| (9) |
First, we examine existence and stability of equilibria of the coupled dynamics
Secondly, let us pay attention to the migration coefficient λ. There are many possibilities with regard to what kind of migration may take place between stocks inside and outside a MPA. The relationship we shall focus on here could allow mutual in- and out-migration. For
Particular cases when
In this setting, one has to deal with the following optimal control problem
| (10) |
3.1 Optimality conditions
The Euler first order optimality condition gives the following equations:
We first study candidate optimal steady state solutions
| (11) |
| (12) |

Graphs of R(⋅) and H(⋅).
Remark
Up to now, there has been no reason to claim that one of the
| (13) |
3.2 Optimal fishing effort and fishery benefit
At optimal steady state, the optimal fishing effort can be derived, combining (8), (9) and (12):
| (14) |
| (15) |
4 Comparison between the two situations
Our goal here is to establish conditions under which a MPA could enhance both economic and the biological situations. We have turned to numerical computation because it seems quite difficult to obtain a complete analytical comparison. In the sequel we shall consider the case where growth functions
We present here some of our numerical results that seem to be particularly relevant for our purpose. For this, we have fixed
4.1 Sensitivity analysis with respect to α and
For different values of α and
|
|
|
|
|
|
|
|
|
| 24 | 0.155 | 0.169 | 0.0262 | 0.241 | 0.462 | 0.111 | 8.6 |
| 30 | 0.137 | 0.172 | 0.0236 | 0.221 | 0.496 | 0.110 | 2.6 |
| 36 | 0.125 | 0.175 | 0.0219 | 0.205 | 0.524 | 0.107 | 1.7 |
| 42 | 0.115 | 0.177 | 0.0204 | 0.193 | 0.550 | 0.106 | 1.3 |
| 48 | 0.107 | 0.178 | 0.0190 | 0.183 | 0.572 | 0.105 | 1.1 |
As expected, we notice that managing a protected area with a higher growth rate (i.e.
4.2 Sensitivity analysis with respect to λ
In practice, it might be difficult to control with accuracy the fish migration between the two areas, and to impose the precise optimal value
The equilibrium
| λ | 7 | 8.6 (optimal) | 10 | |
|
|
|
0.253 | 0.250 | 0.249 |
|
|
0.245 | 0.241 | 0.240 | |
| λ | 1 | 1.1 (optimal) | 2 | |
|
|
|
0.272 | 0.250 | 0.169 |
|
|
0.193 | 0.183 | 0.139 |
We notice that the effects of a variation of the migration coefficient on the steady states densities is not very significant. Consequently, the gain in managing a protected area with a migration coefficient about
4.3 Improvement of the fishery value
Now we compare the present value of the fishery with and without a protected area. More precisely, we consider optimal stationary situations, when the decision to create a reserve or to close the existing one is taken. Therefore the present value of the fishery has to take into account the transit period when the biomass densities have to reach the new steady state. We assume that fishery managers have means to prevent migration outside the MPA when this one has been created (this amounts at taking
Scenario 1
There is no protected area and the value of the stock density is at its optimal value
Scenario 2
There is no protected area and the value of the stock density is at its optimal value
Consider first the durations
When
- (i) Create a protected area at date 0 and prevent fish migration outside the area.
- (ii) Stop the harvest at date
- (i) Stop the harvest at date 0.
- (ii) Create a protected area at date
Scenario 3
The protected area has been created with a migration coefficient
|
|
|
|
|
|
| 24 | 2.3 | 11 | 6.4 | 3.9 |
| 30 | 2.7 | 14 | 7.8 | 4.7 |
| 36 | 3.1 | 17 | 9.4 | 5.4 |
| 42 | 3.4 | 20 | 11 | 6.1 |
| 48 | 3.7 | 22 | 12 | 6.8 |
We check that, in any case, managing a protected area at the steady state
5 Conclusion
In this work, impacts of MPA creation have been investigated, on both economic and biological perspectives. Our attention has been focused on the obtention of theoretical conditions leading to economic benefits on the sole fishing sector. More precisely, it has been assumed that the fishery sector is optimally managed by fishery managers (whether individuals or committees). Their optimal behavior consists then in maximizing the present value of the fishery, defined as the sum of the discounted net revenues derived from the exploitation of the resource. With the help of a two patches model, we have found that MPAs should be installed so that the amount of spillover is maximized. Of course, scientific guides are required to advice fishery managers about the design, location and concrete implementation of MPAs.
In further works, it would be useful to examine conditions under which our results are robust under other management rules. Open access regime should be analyzed first. In the open access case, MPA may act as a management tool amongst other complementary management tools.
Secondly, it is well known that there exist other potential benefits that can be advocated in favor of MPA implementation. It should be actually taken into account of consumer or scientific benefits relative to MPA creation. Considering them together with the fishery profits should allow to define a social value of MPAs. The objective of fishery managers should be then to maximize this social value. In the case where the amount of the resource spillover is not sufficient, MPA may lead to loss for the fishery sector. If managers concerns are the only fishery sector benefits, MPA implementation must be given up. If the objective of fishery managers takes into account others potential benefits, the social value of a MPA may still be positive. This work was obviously beyond the purpose of this paper, and would require to assess both economic and social implications of MPAs. Benefits and costs to extractive users (fishermen) but also benefits and costs to nonextractive users, as well as management benefits and costs should be estimated. (Sanchirico et al. [11]). Moreover social value assessment of the MPA would require to take into account equity issues, which may arise because MPAs affect generally different users groups in a disproportionate way.
Appendix A
We study the equilibria of the coupled dynamics
| (A.1) |
Proposition 1
When the functions
- (P1)
- (P2)
Proof
Consider the two functions on
| (A.2) |
From the properties (P1) and (P2), we deduce that these functions fulfill the following properties:
- (P3) the graph of
- (P4) the symmetric of the graph of
- – the graph of
- – the symmetric of the graph of
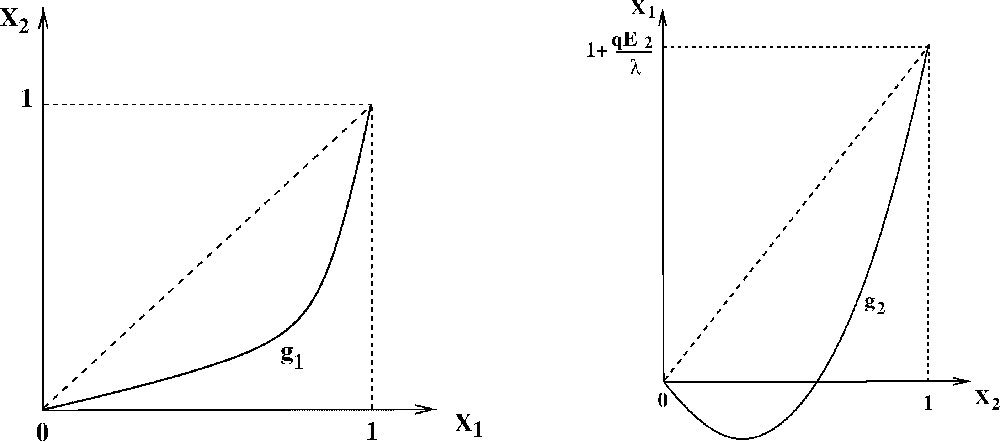
Graphs of the functions
Notice that
| (A.3) |
| (A.4) |
Corollary 2
When
Proof
We have

Existence and uniqueness of
Appendix B
When
Furthermore, by the mean value theorem, there exists an unique
Vous devez vous connecter pour continuer.
S'authentifier