

1 Introduction
Parnassius apollo (Lepidoptera: Papilionidae) has attracted attention of naturalists and insect-collectors for many decades [1–3]. It is one of the biggest butterflies in Europe among the uncommon ones, and its populations are rather locally distributed [4]. The last glaciation in Europe and the subsequent periods caused a selective pressure on the species, which resulted in the appearance of numerous morphologically-distinct local forms, or even of subspecies [2,5,6]. Since their spatial distribution was often limited to a single mountain range, many of the forms have suffered from the changes in the local conditions. Consequently, some of them severely decreased in numbers or even were threatened with extinction [4,7–10].
On the other hand, the morphological variability of the species raised questions concerning its origin. Many authors pointed out the interactions between distribution of the local forms and their food-plants as a possible reason (e.g., [6,11–13]). P. apollo is considered to be a typical stenophagous species feeding only on various Sedum plants and occasionally on Sempervivum [14,15]. European forms of Apollo were divided into two trophic groups according to the preferred food-plants, the first one feeding on elongate, leafy species, and the other preferring creeping forms with tiny roll-like leaves. The first group – ‘telephiophagous’ – includes subspecies feeding predominantly on Sedum telephium and related species, while the second one (‘albophagous’) includes forms feeding on Sedum album [6,16–18]. Previous studies on the Apollo biology in Europe allowed us to depict a distribution map of ‘telephiophagous’ [8,11,12,14,17,19–26] and ‘albophagous’ [8,11,12,14,17,21,23–35] populations (Fig. 1). Approximate boundary between these two groups crosses Central Europe. In the Carpathians, this boundary is more problematic, since ‘telephiophagous’ and ‘albophagous’ forms occupy areas situated very close to each other [17,24,25]. Deschamps-Cottin and co-workers have shown that the apollo larvae from the French Alps did not display strict preferences towards particular Sedum plants and they grew and developed successfully both on Sedum of ‘telephium’ and ‘album’ type. However, having the choice, the larvae always preferred the plant growing naturally in their original biotope [11]. Both forms of P. apollo from the Polish part of Carpathians: ssp. frankenbergeri Slaby, inhabiting the Pieniny Mts, and ssp. niesiolowski, from the Tatra Mts, belong to the ‘telephiophagous’ group; however, literature data point out different food-plants for them (Sedum telephium = S. maximum and S. fabaria, respectively). Some authors suggested an even broader spectrum of food-plants [13,17,18,36]. Field records from the late 1980s in the Polish part of the Pieniny Mts revealed that only about 20 larvae of P. apollo survived in one locality [37]. In order to prevent the subspecies from total extinction, a recovery plan has been launched in 1991, aiming at founding its colonies at sites, where the species was present before 1950 [38]. First and foremost resources of food plant (S. telephium) in the previous apollo biotopes were estimated and locally enlarged [39–41]. After a decade of work, number of individuals inhabiting several sites within the massif is estimated to be about 1000 [41–43]. The success of the program depended significantly on the semi-natural colony of the apollo subspecies, set up close to the Pieniny National Park under the supervision of the scientific board of the park. However, rearing the Apollo larvae in the colony needs providing sufficient amounts of food-plants. That is of particular importance for the last-instar larvae, consuming daily up to 4 g of fresh leaves [44]. Natural resources of S. telephium in the Pieniny Mts, exploited by wild P. apollo individuals, limit further the growth of the Apollo population. There is no special S. telephium plantation for the colony maintenance in the Pieniny Mts, hence larvae are fed on the plants collected at other sites within the Pieniny range and outside it. So, any cost-effective alternative of feeding the colony individuals is desirable. However, orpine sprouts cultivated in glasshouse conditions seem to be unsuitable to meet the feeding demands of the Apollo larvae colony, since their use have caused significant mortality of the caterpillars, probably due to different composition of metabolites in comparison with plants growing in natural conditions [3,34]. In this study, we aimed to define the feeding preferences of P. apollo ssp. frankenbergeri larvae to assess the possibilities of successful rearing of the colony individuals on more than one commercially available Sedum species.
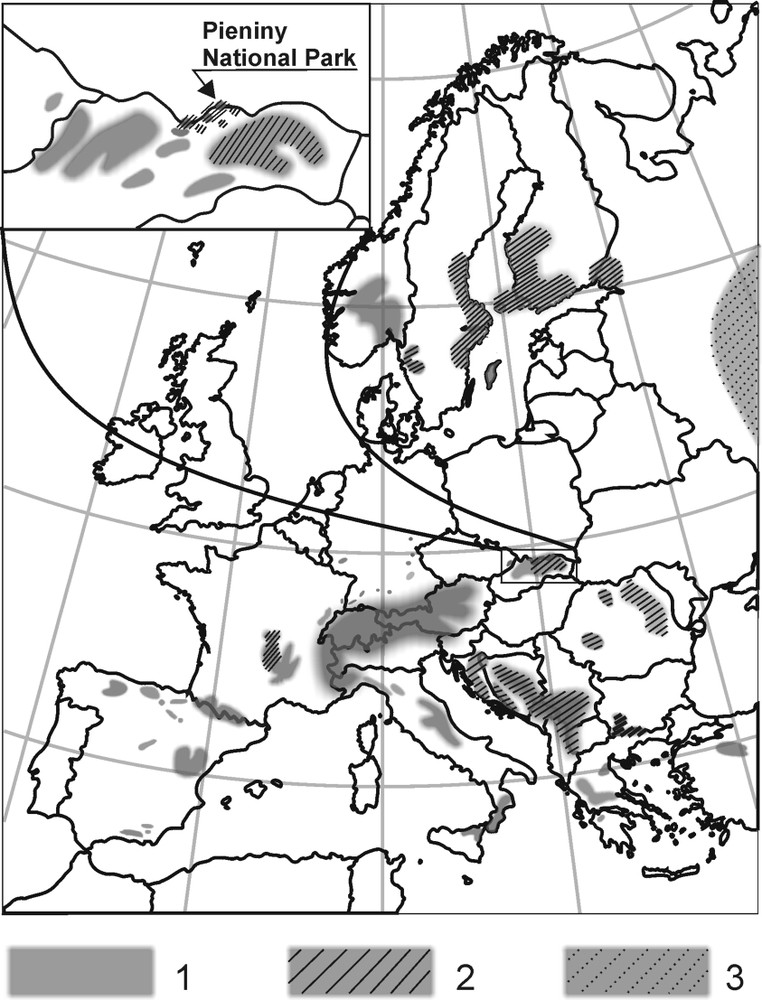
European distribution of main trophic groups of P. apollo. Inset shows approximate areas in Middle Carpathians: (1) ‘albophagous’ populations or subspecies; (2) ‘telephiophagous’ populations or subspecies; (3) contradictory data. Compiled from various authors (see text for references).
2 Materials and methods
Since Apollo is protected by national and international law [4,45,46], necessary permissions for rearing and breeding it were obtained from the Polish Ministry of Environmental Protection and State Nature Conservator. All P. apollo ssp. frankenbergeri feeding preferences tests were carried out on individuals from the semi-natural open-field breeding colony set up for a recovery plan. The colony shares genes pool with the wild population due to the bi-directional flow of genes. Emerging first-instar apollo larvae, after over-wintering in natural conditions, were placed in large glass insectaries () covered by soil layer with various growing plants, including Sedum telephium and Sempervivum sp. Such conditions were suitable and provided young larvae with the appropriate food amounts. Starting from the third instar, freshly collected sprouts of S. telephium were provided as food. Larvae were exposed to natural weather conditions. Only in case of heavy rains, the insectaries were covered with glass lids to avoid larvae drowning. In these conditions, larvae developed even slightly faster than in their natural biotopes, probably due to more stabilised thermal conditions. For feeding preferences tests, all commercially available cultivated Sedum species were purchased. These were the following: S. floriferum, S. ewersii, S. kamtschaticum, S. spectabile (species with flattened leaves); S. album, S. reflexum, S. rupestre, S. rosea and S. spurium (species with terate leaves). Sedum taxonomy is still problematic, so the names given by the purveyor were confirmed according to Flora Europea and Royal Botanic Garden Edinburgh (RBGE) database (Table 1) [47,48]. The former source was also used to unify nomenclature of ‘telephiophagous’ apollo subspecies food-plant, since earlier authors often used various Latin names (Table 2) [48]. All purchased plants grew in natural weather conditions, because previous studies had shown that the plants with artificially accelerated growth (glasshouse cultivation) were unsuitable as the food-source for the Apollo larvae [3,34]. Potted plants were transported to the Apollo colony in the Pieniny Mts and placed in a separate insectary for acclimation. The main experiment was preceded by a preliminary test, in which samples of all the plants were offered to the group of fourth instar (L4) Apollo larvae. This preference test allowed selection of edible plant species for further experiment, carried out on the last-instar (L5) larvae. To assess the effects of selected plants on larval vitality and further development to adult stage, late fourth-instar larvae similar in size and weight were selected from the colony. At the end of April, three experimental groups (I–III; 15 individuals in each) were set up on different menu options. Larvae of group I were fed on Sedum kamtschaticum var. floriferum and S. rupestre, larvae of group II on S. album and S. ewersii, and the larvae of group III (being the reference group) on S. telephium ssp. maximum (the same as that provided to the stock colony insects) only. The larvae from the onset of the last instar were provided every day with fresh Sedum leaves and the remnants were removed. Fresh weight of larvae in each experimental group was recorded until the insects began building the cocoon shell. The measurements as well as observations of development were carried out in 1–3 day intervals, depending on the weather conditions, except for one rainy episode, when larvae almost ceased feeding for 4 days and were almost inactive. To minimise the stress caused by the manipulation, the larvae were quickly weighed without anaesthesia. The dimensions of stretched forewings were measured in the newly emerged adults. Fecundity was estimated by collecting and counting the eggs laid by survived females. Obtained results were analysed by ANOVA/MANOVA with STATISTICA 5.0® package for PC. Post hoc T Tukey test was applied to estimate significant differences among experimental groups ().
Nomenclature of cultivated stonecrops, used in the feeding preferences experiment, according to Flora Europaea and Royal Botanic Garden Edinburgh (RBGE) database. Names used in the text are bold-typed
| Purchased plant | Flora Europaea | RBGE database |
| Sedum album | Sedum album L. | Sedum album L. |
| S. ewersii | S. ewersii Ledeb. | Hylotelephium ewersii (Ledeb.) H.Ohba |
| S. floriferum | – | S. kamtschaticum Fisch. & C.A.Mey. var. floriferum ‘Weihstephaner Gold’ |
| S. kamtschaticum | – | S. kamtschaticum Fisch. & C.A.Mey. |
| S. spectabile | – | S. spectabile Boreau |
| S. reflexum | S. rupestre L. | S. rupestre L. |
| S. rosea | Rhodiola rosea L. | Rhodiola rosea L. |
| S. rupestre | S. rupestre L. | S. rupestre L. |
| S. spurium | S. spurium Bieb. | S. spurium M.Bieb. |
Names of food-plants of ‘telephiophagous’ apollo subspecies, used by earlier authors (‘original name’) and corresponding nomenclature, according to Flora Europaea and the Royal Botanic Garden Edinburgh (RBGE) database. Names used in the text are bold-typed
| Original name | Flora Europaea | RBGE |
| Sedum maximum (L.) Suter | S. telephium L. ssp. maximum (L.) Krocker | Hylotelephium telephium (L.) H.Ohba ssp. maximum (L.) Ohba |
| S. telephium L. | S. telephium L. ssp. telephium | H. telephium (L.) H.Ohba |
| S. fabaria Koch | S. telephium L. ssp. fabaria (Koch) Kirschleger | H. telephium (L.) H.Ohba ssp. fabaria (Koch) H.Ohba |
| S. purpureum Schultes | S. telephium L. ssp. telephium | H. telephium (L.) H.Ohba ssp. telephium |
| S. carpaticum G.Reuss | S. telephium L. ssp. fabaria (Koch) Kirschleger | – |
3 Results
The preliminary test confirmed that food plants spectrum of P. apollo larvae inhabiting the Pieniny Mts is narrow. Sedum kamtschaticum, S. spectabile, S. rupestre, S. spurium and Rhodiola rosea were even not bitten, although caterpillars actively searched for food. The main experiment revealed that successful development of the last-instar larvae until pupation depended strongly on the plants available as food. Only larvae fed on S. telephium ssp. maximum all developed into pupa. Average duration time of the L5 instar was also the shortest (18 days) in this group (Figs. 2 and 5). Feeding on other Sedum species increased L5 life span by about two weeks. Moreover, a high mortality was stated – 60% in group I and 87% in group II (Figs. 3 and 4). It should be noted that larvae fed exclusively on S. kamtschaticum var. floriferum and S. rupestre, almost did not develop. Hence, pupation of the larvae in this group took place only due to including, after 12 days of experiment, S. telephium ssp. maximum, as an additional food source (Fig. 2).
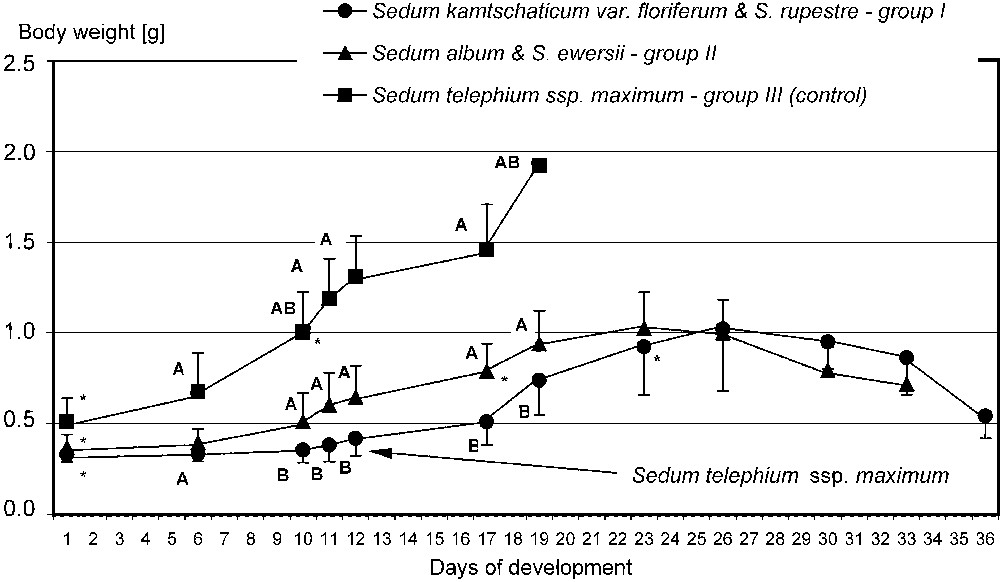
Changes in fresh body weight of L5 apollo larvae fed on different Sedum species. The same letter (A, B) indicates a significant difference between the groups for a particular day of measurements; the asterisk (∗) indicates the day in which the larva body weight in the group was significantly higher than at the beginning of the experiment.
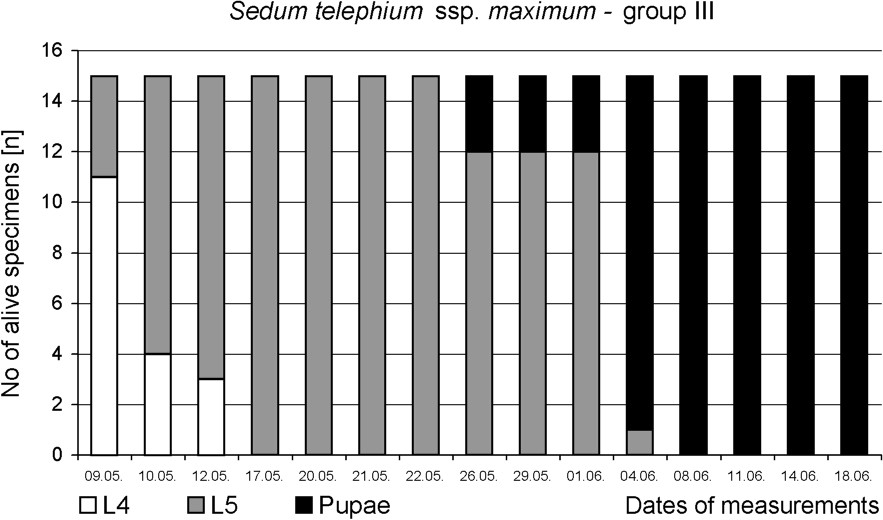
Survival and development until pupa of P. apollo fourth-instar larvae fed on S. telephium ssp. maximum.
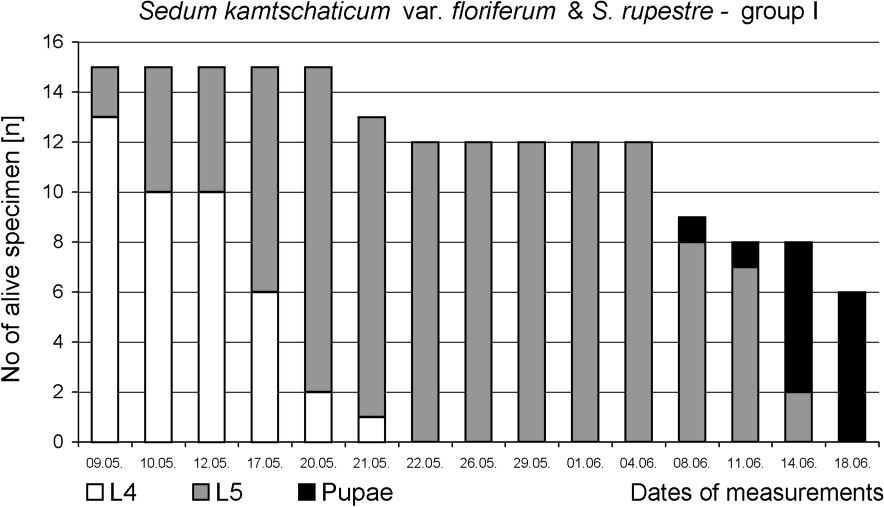
Survival and development until pupa of P. apollo fourth instar larvae fed on S. kamtschaticum var. floriferum and S. rupestre (+ S. telephium ssp. maximum).
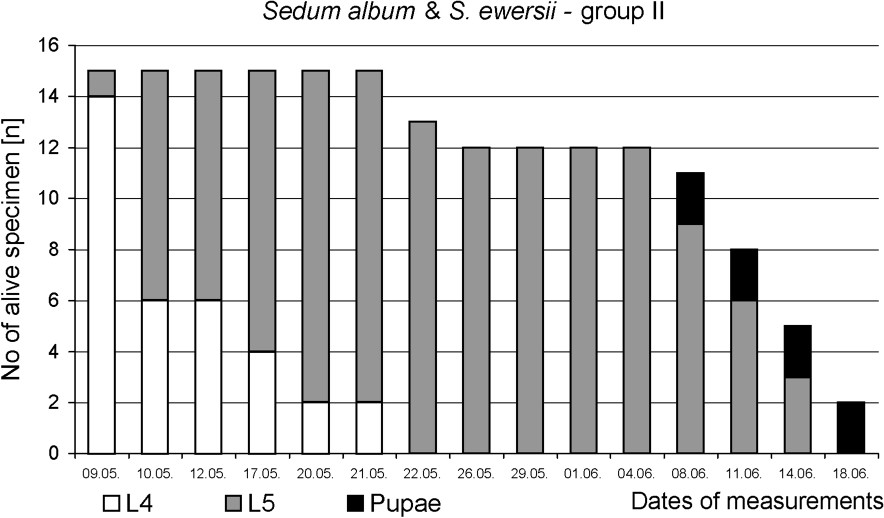
Survival and development until pupa of P. apollo fourth-instar larvae fed on S. album and S. ewersii.
The fastest growth of the larvae was observed in group fed on S. telephium. After nine days, L5 larvae doubled their initial fresh body weight (0.391 g), and during following days they reached 1.458 g on average (Fig. 2). The maximum individual fresh body weight was 1.925 g. Larvae fed S. album and S. ewersii needed almost three weeks to double their initial fresh body weight. Despite further slow growth only two heaviest individuals in the group pupated (Figs. 2 and 4). In group I, young L5 larvae tasted the offered plants but wandered all over the insectary in search for other food. Before S. telephium was added, body mass increased only by 27%. Despite further significant growth on S. telephium 50% mortality had been observed during pupation, mainly among larvae with low body weights (Figs. 2 and 3). One day before they started to produce cocoon, larvae from group I and II had also significantly lower fresh body weights compared with those of group III (Fig. 6). Decreased consumption of food in the last larval instar affected consecutive developmental stages. One of effect was increased pupal mortality – 55.6% in group I and 50.0% in group II, compared with 13.0% in the control group. Adults that emerged from pupae in group I and II were also smaller than the individuals fed on S. telephium leaves. The latter had significantly longer and broader forewings. The smallest wings occurred in adults from the group fed on S. album and S. ewersii (Fig. 7). Unsuitable food offered to larvae during the last instar resulted also in small number of laid eggs (Table 3).
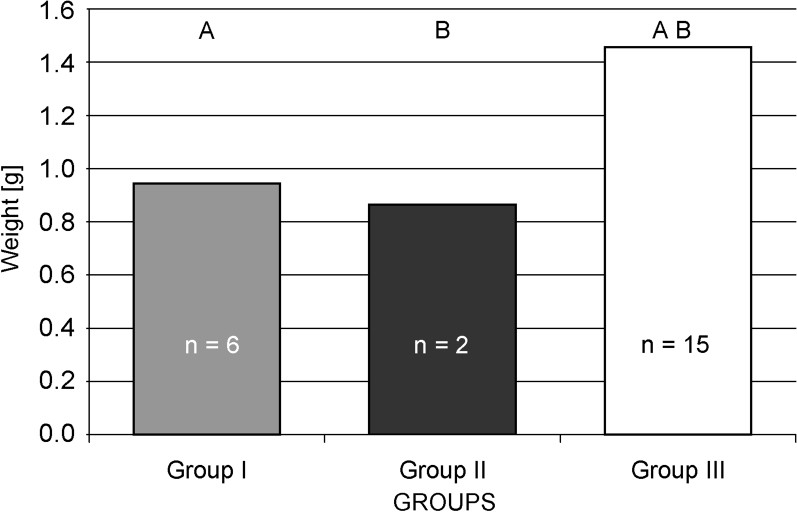
Mean P. apollo prepupae weight in the experimental groups.
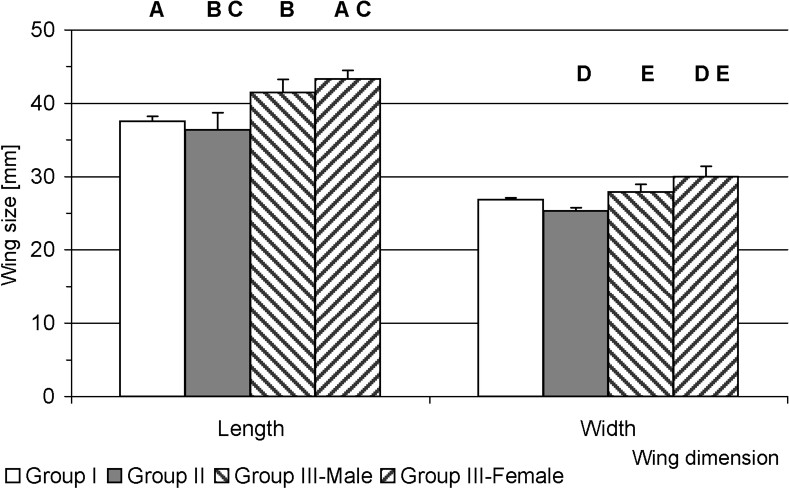
Forewing size (mm) in apollo adults that emerged from pupae in experimental groups. The same letter indicates significant differences between the groups.
Adult fecundity (number of eggs per female) in the experimental groups
| Group | Eggs/female | |
| I | (S. kamtschaticum...) n=1 | 14 |
| II | (S. album...) n=1 | 26 |
| III | (S. telephium...) n=5 | 68.4 |
4 Discussion
Three plants species among those offered to Apollo caterpillars S. telephium, S. spectabile and S. ewersii represented the Hylotelephium group. Closely-related S. telephium ssp. fabaria, recorded as a food plant of P. apollo ssp. niesiolowskii, inhabiting the northern part of the High Tatra range [25], rarely occurs in the Pieniny range and in its vicinities. As it was also unavailable commercially, it was not included in the study. Among European Sedum representatives there were S. rupestre, S. album and S. spurium, of which S. album has widest distribution range in Europe, grows at very contrasting altitudes and in a great variety of soils [49]. Sedum kamtschaticum and its variety floriferum, represented far eastern, and Rhodiola rosea, inhabiting the European mountain ranges are circumpolar stonecrops species. Apollo larvae inhabiting the Pieniny Mts grow and develop successfully only when they have sufficient amounts of S. telephium ssp. maximum. Slower and prolonged larval development on S. album and its almost complete inhibition on S. kamtschaticum var. floriferum and S. rupestre, led to high larval and pupal mortality and low fecundity of the adults. These findings as well as the rejection of the eastern Asiatic S. spectabile point out that this Apollo subspecies belongs definitely to ‘telephiophagous’ group, and has strict food preferences. Deschamps-Cottin and co-workers [11] discovered that Apollo strains, originating from various regions, developed into healthy adults both on S. album and on S. telephium, although the individuals fed on S. telephium were generally larger. Moreover, they have shown that fourth- and fifth-instar apollo larvae collected at various localities in France and Sweden attacked sprouts of almost all (7) offered Sedum, including S. altissimum, and S. ochroleucum, as well as Sempervivum arachnoideum and S. montanum. It is noteworthy that neither S. altissimum nor S. ochroleucum was previously cited among food-plants of P. apollo. Vegetative forms of these European species are difficult to distinguish between each other and from S. rupestre, used in the present study [49]. These findings led French authors to conclude that Apollo larvae are oligophagous, depending on the plants available in the feeding place. However, they also observed that larvae fed on S. ochroleucum developed much longer and slower and finally all individuals died [11]. This coincides with our findings on S. rupestre, which, together with S. kamtschaticum var. floriferum appeared unsuitable food source for the Apollo larvae. In the French study [11], Apollo strains from different origin showed various preferences toward Sedum and Sempervivum plants. The broadest spectrum had strains from the Gotland Island and the Southern Alps near Briançon, while strains from the French Massif Central were the most selective [11]. Apollo larvae from the Pieniny Mts appear to be even more specific. A question that remains is whether they could use S. telephium ssp. fabaria as a food source, but certainly S. album or S. ewersii are much less suitable than S. telephium. Our results give further evidence that P. apollo in Europe falls into ‘telephiophagous’ and ‘albophagous’ trophic groups. This division probably reflects two main routes of its westward migration from Central Asia in the Tertiary and subsequent changes in distribution following glacial and postglacial periods in Europe [6,17,25], with S. telephium being its prime food-plant. Populations that colonised mountain ranges in western Europe had to change its feeding into the available ‘lower’-quality food, and became ‘albophagous’. This explains why they still develop better and grow larger on S. telephium [11,34], while ‘telephiophagous’ forms cope much worse on S. album [17]. This raises a question about the reason of such ‘trophic asymmetry’ among these groups. It has been suggested that the larval performance may depend on the composition of secondary metabolites in their host plant [11,34]. Phytochemical data point out that S. album and S. telephium contain different amounts and composition of phenolic and flavonoid compounds [50–53]. Effect of a specific set of these allelochemicals on Apollo larvae performance and development remains so far unexplained. It may be concluded, that albophagous strains or subspecies of P. apollo from mountain ranges in Southern and Western Europe remain oligophagous, but telephiophagous forms from central Europe should be considered as monophages.
Acknowledgments
We express our thanks to Managing Director and Scientific Board of The Pieniny National Park for administrative support and Mr Tadeusz Oleś for his expertise and help in Apollo breeding.