

1 Introduction
Until about two decades ago, the number of scorpion family-groups had been stable for almost a century [1]. Subsequent modifications of the familial classification of scorpions resulted mainly from revision work in which subfamilies were raised to the family rank [2]. In very few cases, the descriptions of new families resulted from entirely original discoveries [3], but even in these cases, the results did not always reach a consensus among experts [4].
A recent remarkable discovery was that of Pseudochactas ovchinnikovi Gromov, 1998 in an isolated mountainous region of southeastern Uzbekistan and southwestern Tajikistan, in Central Asia. In fact, this scorpion displays several characters that are unique among recent (extant) scorpions, including a distinct trichobothrial pattern and a number of other characters. Besides the combination of features shared with buthid and nonbuthid scorpions, Gromov [5] noted several unique characters of Pseudochactas that warranted the creation of a new monotypic family, Pseudochactidae Gromov, 1998. Subsequent authors have not reached a consensus regarding the phylogenetic position of this enigmatic scorpion. Based on the peculiar trichobothrial pattern of Pseudochactas, Fet [6] suggested a relationship to the most plesiomorphic Buthidae C.L. Koch, 1837 or to Chaerilidae Pocock, 1893. Lourenço [2] placed Pseudochactas in a new superfamily, Chaeriloidea Pocock, 1893, implying that he considered it to be the sister group of Chaerilus. Although there is widespread agreement that Pseudochactas is basal within recent scorpions, its precise phylogenetic position remains a matter of debate [7]. In an exhaustive study of P. ovchinnikovi, Prendini et al. [8] concluded that the most plausible position for this ‘living fossil’ should be a sister group of Buthidae.
Another controversial aspect surrounding the pseudochactid lineage concerns its period and region of origin. Fet et al. [7] speculated about a possible Pangaean origin (Permian–Triassic), whereas Prendini et al. [8] estimate that given the family's restricted geographical range and basal phylogenetic position, it should be considered as a palaeoendemic, a relict of an earlier scorpion fauna living in a more mesic environment.
The present discovery of a second element belonging to the pseudochactid lineage in Laotian limestone caves reopens this question (Fig. 1).

Map showing the type locality of Troglokhammouanus steineri sp.n., in Laos.
2 Orogeny and geodynamics of Laos and ecology of the Hin Namno area
The Indochina plate forms the core of the geological structure of southeastern Asia. This plate comprises the countries of Vietnam, Laos, Cambodia and western Thailand, but according to Metcalfe [9], also the southeastern portion of the Malayan Peninsula, a fragment of Sumatra, and the westernmost portion of Borneo.
The Southeast Asia plate originated during the Proterozoic as a portion of Gondwana. It was detached during the Palaeozoic and drifted northward. The carbonate platforms were developed during the Devonian–Late Palaeozoic. The Palaeozoic history of detachment and collision is quite speculative. The equivalent of the Caledonian orogeny followed by the formation of the Palaeotethys Ocean is quite possible. Climate records indicate major differences between Sibumasu, Indochina, and South China during the Late Palaeozoic. During Triassic time, as a result of the Indosinian orogeny and closure of the Palaeotethys Ocean, the Southeast Asian plate joined the Asian continent [9–11].
According to Senghor and Hsü [10], the present region of Uzbekistan/Tajikistan was most probably located on the old Asian core zone, whereas Laos would be composed in part of the old Asian core, but also of several plates that originated as parts of Gondwana, detached subsequently and drifted northward. Although the Palaeozoic history of detachment and collision is quite speculative, these events took place before the Cretaceous. Consequently, the only possible connection between Uzbekistan/Tajikistan and Laos is the old Asian core.
Tham Xe Bangfai is one of the largest river caves in the world (Fig. 2). The trunk passage is a large tunnel 40 to 80 m wide and high, becoming larger in some zones. In many portions of the cave, it is completely filled with water during the dry season. There are a few sections, particularly higher passages, with sand, gravel, or rock banks with rock and stall walls (Fig. 3). The underground river consists of calm stretches, without a notable current, separated by six different rapids. The water is clear and deep. The altitude at the river level is ca. 200 m a.s.l., while that of the surrounding mountains reaches 600 m a.s.l. Temperature in the cave is 20 °C on average, while humidity reaches 90%.


Tham Xe Bangfai cave in the Province of Khammouan. 2. External view of the resurgence side (Photo by H. Steiner). 3. ‘Internal beach’, where the holotype was collected (photo by C. Ghommid).
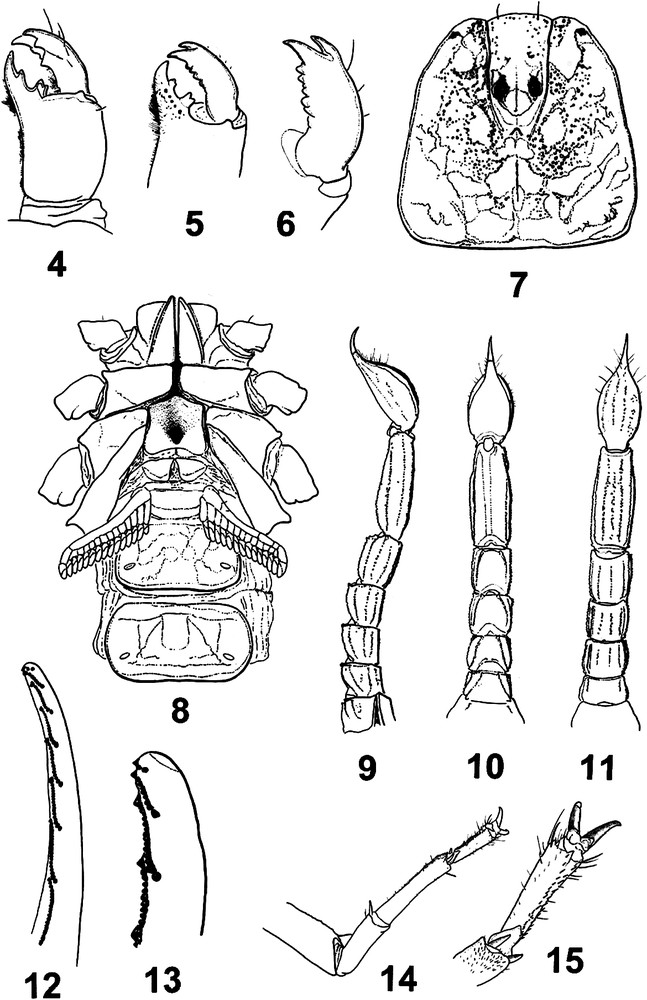
Troglokhammouanus steineri sp.n., female holotype. 4–6. Chelicerae, dorsal and ventral aspects, and detail of movable finger, ventral aspect. 7. Carapace. 8. Ventral aspect, showing coxapophysis, sternum, genital operculum, pectines, and sternites III–IV. 9–11. Metasoma and telson, lateral, dorsal and ventral aspects. 12. Movable finger of pedipalp chela with subrows of granules. 13. Extremity of the finger in detail. 14. Leg IV, showing spurs. 15. Detail of telotarsi with a pair of ventrosubmedian rows of spinules.
A large colony of swiftlets was observed in the entrance hall (resurgence side), and bat colonies deeper into the cave. Over the whole length of the 7-km underground river, many insects are attracted to artificial light, mainly small beetles, flies, mosquitoes, and mayflies. Scorpions were collected about 2 to 3 km from the entrance on alluvial soil and on a sand bank next to the edge of the river. Collections in the same site also included several insects, spiders, opilions, isopods, and millipedes.
Hin Namno is an area where the Central Indochina Limestone meets the Annamite Chain. As a result, this area has prominent limestone escarpments and many caves, including the system along the Xe Bangfai River [12]. The specific zone where the caves are located is placed in the northern Annamite ecoregion [12]. The area exhibits a mosaic of semi-evergreen and mixed deciduous forests, whilst pockets of closed canopy evergreen forest are found within the karstic zones. Further, there are areas of secondary forest, shifting cultivation and bamboo forest. Due to difficult access, much of the forest of Hin Namno is free from extensive exploitation, with the exception of areas near roads and villages.
3 New biogeographical conjectures
In their respective studies of Pseudochactas ovchinnikovi, Fet et al. [7] and Prendini et al. [8] insisted about the restricted distribution of this monotypic family to the mountains of Uzbekistan and Tajikistan. According to Fet et al. [7], the relict character of this taxon's distribution could be due to its perseverance in mild-climate, low-mountain depressions, with surrounding lower altitude deserts. This opinion is partially shared by Prendini et al. [8]. Furthermore, Fet et al. [7] suggested that based on its ancestral position in scorpion phylogeny, the Pseudochactidae lineage evolved during the Permian/Triassic. They assumed, however, that it was impossible to speculate on whether this lineage was localized or widespread since there are no fossils belonging to Pseudochactidae, and the family is represented by a single monotypic genus. Although several key aspects are unknown to reconstruct properly the biogeographic origins of this group, Soleglad and Fet [13] and Fet et al. [7] discussed the possible Pangaean origin (Permian to Triassic time) of four scorpion lineages, including the Pseudochactidae. Again, according to Fet et al. [7], through the Cretaceous, many animal taxa persisted as relicts, while other perished during global climatic changes and associated restructuring of ecosystems [14]. The Pseudochactidae, the most primitive extant group of scorpions (sensu Fet et al. [7]), appears to have survived in such relict conditions. Hence, given this scenario, the Pseudochactidae should have split from all other scorpion lineages well before the Triassic [7,13].
Prendini et al. [8] largely rejected the conclusions of Fet et al. [7] on the temporal origins of this family. Rather, they considered that given the restricted geographical range and basal phylogenetic position of Pseudochactas, it should be considered a palaeoendemic, a relict of an earlier, probably mesic environment scorpion fauna that probably existed in the region by Upper Cretaceous and Lower Tertiary times, before climate change eliminated other members of its lineage.
Prendini et al. [8] stated that the conclusions by Soleglad and Fet [13] and Fet et al. [7], extrapolating that Pseudochactas diverged from all other scorpion lineages ‘well before the Triassic’ (more than 250 Ma), rest solely on the confirmation of its phylogenetic position as the sister group of all other recent scorpions. Prendini et al. [8] conclude, however, that the most plausible hypothesis is to consider Pseudochactas as the sister group of Buthidae. Consequently, its divergence could be more recent. They insisted also on the fact that a more recent divergence accords better with the suggestions by Soleglad and Fet [13] and Fet et al. [7] that Pseudochactas may represent a remnant of a littoral or insular fauna of the Tethys Sea, that evolved towards the end of the Cretaceous and became elevated by mountain uplift during the Tertiary, hence, surviving the onset of aridification in Central Asia.
The recent discovery of a new genus and species of Pseudochactidae in a Laotian cave system re-opens the question about the palaeo-biogeographic origin of this lineage. As stated in the orogeny section, the only possible land connection between Uzbekistan/Tajikistan and Laos is the old Asian core. Consequently, the present known geographic disjunction in members of this family reflects a much larger past geographic area of the lineage, and the hypothesis of a possible Pangaean origin (Permian to Triassic time) needs to be reconsidered.
4 Taxonomic treatment
Family Pseudochactidae Gromov, 1998
Genus Troglokhammouanus gen. n.
Diagnosis. Cheliceral movable finger, dorsal edge with three teeth (medial, subdistal, external distal); external distal tooth smaller than internal distal tooth. Anterior margin of carapace with a moderate convexity, posterior margin shallowly recurved. One pair of very small lateral ocelli. One pair of circumocular sutures with a broad U-shaped configuration, but incomplete in the posterior region to the median ocular tubercle. Anterosubmedian carinae absent from the zone internal to the circumocular sutures. Type-D trichobothrial pattern [13,15] with 35 trichobothria per pedipalp: 12 on the femur (six dorsal, three internal, three external), 10 on the patella (three dorsal, one internal, six external, trichobothria absent on the ventral surface), and 13 on the chela (five manus, eight fixed finger); pedipalp femur dorsal trichobothria with beta (β) configuration. Pectinal peg sensillae (in study). Sternum pentagonal, type 1 [16], moderately compressed horizontally, slightly longer than wide, external aspect not flat, with a concave region, posteromedian depression round. Telotarsi each with a pair of ventrosubmedian spinule rows. Metasomal segment V with a well-developed pair of ventrosubmedian carinae, and one well-developed ventromedian carina between ventrosubmedian carinae, continuous throughout the length of segment. Fixed and movable fingers, dentate margins each with median denticle row comprising 7/8 oblique granular subrows. Respiratory spiracles small, oval in shape; tibial spurs present on legs III and IV; pro- and retrolateral pedal spurs present on legs I–IV.
Derivatio nominis. The generic name is a combination of Troglo (from troglobitic = cave living) and khammouanus (from the Province of Khammouan). Type species Troglokhammouanus steineri sp.n.
Troglokhammouanus steineri sp.n. (Figs. 3 and 4 (4–24))
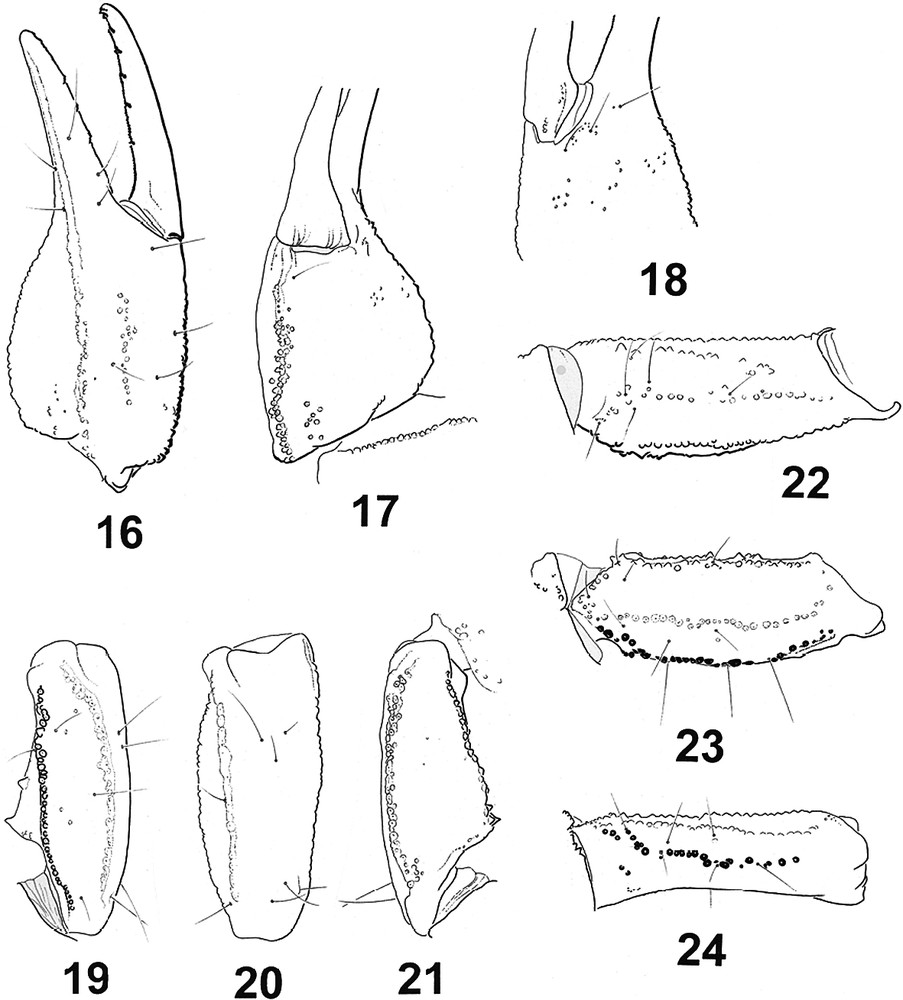
Troglokhammouanus steineri sp.n., female holotype. Trichobothrial pattern. 16–18. Chela, dorso-external, ventral and internal aspects. 19–21. Patella, dorsal, external, and ventral aspects. 22–24. Femur, internal, dorsal and external aspects.
Diagnosis. As for the new genus.
Type material. Female holotype; male (juvenile) paratype (prepared for SEM studies), Laos, Province of Khammouan (Fig. 1), Boualapha, Tham Xe Bangfai cave ( N– E), mid-section of cave, 14/II/2007 (H. Steiner). Types should be deposited in the collection of the ‘Muséum national d'histoire naturelle’, Paris.
Patronym. In honour of Dr Hemut Steiner, Hanau, Germany, who collected the specimens.
Description based on the female holotype (measurements are given in millimetres after the description).
Colour. General coloration reddish–yellow to yellowish–brown; carapace and tergites with very diffused brownish spots; pedipalps reddish–yellow with dark zones over carinae; metasoma reddish–yellow; telson paler than metasomal segment V; chelicerae, legs, coxapophysis, sternum and genital operculum yellowish; pectines pale yellow.
Morphology. Chelicerae: fixed finger, dorsal edge with four teeth (basal, medial, subdistal, distal); ventral edge with 5–6 denticles; movable finger, dorsal edge with three teeth (medial, subdistal, external distal), without basal teeth; ventral edge with 5–6 denticles; external distal tooth smaller than internal distal tooth; ventral aspect of fingers and manus with numerous macrosetae. Carapace: anterior margin convex posterior margin shallowly recurved, almost straight. One pair of very small lateral ocelli. Median ocular tubercle situated anteromedially, comprising pair of median ocelli, larger than lateral ocelli, with pair of weak, smooth superciliary carinae; interocular furrow obsolete. One pair of circumocular sutures with a broad U-shaped configuration, but incomplete in the posterior region to median ocular tubercle. Anteromedian furrow shallow; posteromedian furrow shallow anteriorly, becoming slightly deeper posteriorly; posterolateral furrow shallow, weakly curved; posteromarginal furrow narrow, very shallow. Carapacial surface moderately to strongly granulated, except on lateral and posterior margins; acarinate; anterosubmedial carinae absent from the zone internal to the circumocular sutures. Pedipalps: segments almost apilose, sparsely covered in short microsetae and occasional macrosetae. Femur with five discernable carinae; internomedian carina weak; dorsoexternal, dorsointernal, and ventrointemal carinae well developed; externomedian carina moderate; ventromedian carina vestigial; intercarinal surfaces smooth. Patella with six discernable carinae; dorsoexternal, ventroexternal and ventrointernal carinae well developed; dorsointernal carina well developed, granular; anterior process well developed, comprising a pair of dorsal and ventral spinoid tubercles, the dorsal more strongly developed; externomedian carina absent; intercarinal surfaces smooth. Chela with 6–7 discernable carinae; dorsomedian carina absent; digital carina well developed; ventroexternal carina well developed; ventromedian carina obsolete, reduced to vestigial granules proximally; ventrointernal carina also obsolete, reduced to a few isolated granules; internomedian and dorsointernal carinae weakly developed, each comprising a series of isolated granules; intercarinal surfaces smooth except for coarse, scattered granules on internal surface of manus. Fixed and movable fingers, dentate margins each with median denticle row comprising 7–8 oblique granular subrows; each subrow comprising several small granules and a large proximal granule. Trichobothria: Orthobothriotaxic, type D [13,15], β (beta) configuration, d2 situated on dorsal surface, d3 and d4 in same axis, parallel and closer to dorsoexternal carina than d1, angle formed by d1, d3 and d4 opening toward internal surface; totals: femur, 12 (six dorsal, three internal, three external); patella, 10 (three dorsal, one internal, six external); chela, 13 (five manus, eight fixed finger). Legs I and II, tibiae, without spurs; III and IV, with spurs. Basitarsi each with a pair of pro- and retrolateral pedal spurs. Telotarsi each with a pair of ventrosubmedian rows of spinules. Sternum, pentagonal, type 1 [16], moderately compressed horizontally, slightly longer than wide, external aspect not flat, with a concave region, posteromedian depression round. Pectines: each pecten with three distinct marginal lamellae present in female, 13–14 well-delineated median lamellae present in female. Fulcra present but reduced. Pectinal tooth count: 14–14/14–15 (male and female). Pectinal peg sensillae (in study). Genital operculum: Operculum completely divided longitudinally. Genital plugs not clearly observed in female. Mesosoma: Pre-tergites smooth and shiny. Posttergites II–VI moderately to strongly granular, granulation becoming slightly coarser near posterior edges; VII with a thin granulation and a pair of costate granular dorsosubmedian and dorsolateral carinae, reaching the posterior edge of segment. Sternites almost entirely smooth, acarinate; surfaces with scattered macrosetae; distal margins each with sparse row of macrosetae; respiratory spiracles small, oval in shape. Metasoma almost apilose, very sparsely covered in short microsetae. Eight carinae on segment I, ten carinae on segments II–IV, nine on segment V. Dorsosubmedian carinae well developed, costate granular throughout the length of segments I–IV, absent on segment V; each terminating distally with a enlarged spinoid granule on segments I–IV. Dorsolateral carinae well developed, costate granular throughout the length of segments I–V. Median lateral carinae well developed in segments I–V. Ventrolateral carinae well developed, but becoming obsolete proximally on segments I–II, continuous throughout the length of segments III–V. Ventrosubmedian carinae absent on segment I, weak on segment II, well developed, granular throughout the length of segments III–V. Ventromedian carina on segment V moderately to strongly marked between ventrosubmedian carinae. Intercarinal surfaces smooth. Telson vesicle smooth dorsally, with moderately marked granules laterally and ventrally; aculeus without a subaculear tubercle ventrally. Venom glands unknown.
Geographic distribution. Only known from the type locality (Fig. 1).
Morphometric values (in mm) of the female holotype. Total length 39.1. Carapace: length 5.3; anterior width 3.0; posterior width 5.4. Mesosoma length 11.2. Metasomal segments: I, length 2.0, width 2.9; II, length 2.2, width 2.6; III, length 2.5, width 2.5; IV, length 3.1, width 2.3; V, length 5.9, width 2.3, depth 2.1. Telson length 6.9; vesicle length 4.9, width 2.6, depth 2.3. Pedipalp: femur length 5.1, width 2.0; patella length 5.5, width 2.1; chela length 8.9, width 3.3, depth 2.6; movable finger length 5.1.
Acknowledgements
I am most grateful to Bernard Duhem (MNHN, Paris) for preparing the illustrations, to Jean-Claude Rage (MNHN, Paris) for several information on the geology and orogenesis of the Laos region and for the revision of different sections of this paper, and, finally to Steven M. Goodman (Field Museum, Chicago) for the general review of the manuscript.