

1 Introduction
La loi de programme no 2006-450 du 18 avril 2006 pour la recherche, née à la suite du mouvement des chercheurs de 2003, a profondément réorganisé la recherche en France. Un élément de cette loi concerne la création de réseaux thématiques de recherche avancée, matérialisée par la mise en place des instituts du Centre national de la recherche scientifique (CNRS), auxquels appartient l’Institut national de l’écologie et l’environnement (INEE-2009). « Lieu de pluri- et d’interdisciplinarité, l’INEE a vocation à faire émerger les sciences de l’environnement en tant que champ scientifique intégré.1 ». Parmi les instruments incitatifs de l’INEE, figurent les financements de groupement de recherche (GDR), outils de regroupement des chercheurs membres d’unités CNRS ou d’autres tutelles autour d’une thématique précise. Ainsi, ces GDR structurent thématiquement la communauté et viennent se superposer au maillage géographique des unités de recherche classiques. De façon plus générale, ces GDR ont pour objectifs de recenser les ressources à l’échelle nationale sur un champ thématique et de dépasser les clivages disciplinaires. Dans ce contexte, l’INEE a récemment identifié l’écologie trophique comme un de ses champs prioritaires.
La description des relations trophiques entre organismes, c’est-à-dire liées à leur alimentation et à leur nutrition, est une problématique traditionnelle en écologie, aussi bien pour décrire les flux d’énergie dans les écosystèmes [1] que pour comprendre les interactions entre espèces [2]. Toutefois, les développements récents des études du fonctionnement des écosystèmes et l’émergence de nouvelles méthodes d’analyse des relations trophiques ont conduit à la constitution d’un champ original de recherche, l’écologie trophique. Ce pan de recherche vise à comprendre les déterminismes et les conséquences écologiques des relations alimentaires et de l’acquisition des ressources nutritives par les êtres vivants. Cependant, l’INEE souligne que l’hétérogénéité des recherches menées en écologie trophique en France constitue un frein à l’avancement de la discipline (cf. Prospective « Écologie fonctionnelle », [3]). C’est la raison pour laquelle, dès 2013, des subventions accordées par l’INEE et l’Inra ont permis la nucléation, sous l’appellation GRET (Groupe de recherche en écologie trophique, http://gdr-gret.univ-bpclermont.fr/), d’un réseau scientifique rassemblant, au niveau national, les chercheurs et enseignants-chercheurs travaillant sur les réseaux trophiques marins, dulçaquicoles et terrestres. Ce réseau scientifique GRET a montré l’opportunité et les atouts d’une nouvelle structuration de la diversité des recherches en écologie trophique et a officiellement été labellisé GDR CNRS 3716 par l’INEE en janvier 2015.
Les créateurs du réseau ont relevé ce même constat d’hétérogénéité des recherches menées en écologie trophique, qui se manifeste par la diversité des écosystèmes et des organismes modèles, la multiplicité des méthodes et des questions abordées, et un relatif isolement des chercheurs et unités de recherche. Ils ont donc inscrit dans la lettre de mission du GRET l’objectif de visualiser plus clairement les motifs et raisons d’un tel morcellement du paysage français de l’écologie trophique. Le manque de structuration de cette communauté pourrait être d’ordre culturel, dû à un manque d’identification de cette communauté même (sait-on ce qu’est l’écologie trophique ?) et au poids d’un héritage (les scientifiques sont liés à leur écosystème d’étude et aux problématiques habituellement développées dans ces environnements, explorant indépendamment les versants différents des grands concepts et des questionnements écologiques). Selon cette hypothèse, les outils de communication et de réseautage tels que le GRET pourraient prétendre lisser cette hétérogénéité. Une hypothèse alternative serait que les enjeux socio-écologiques sont très fortement liés à la nature des écosystèmes (marins, prairiaux, dulçaquicoles…), dictant ainsi des questions de recherche fondamentalement différentes entre grands types d’écosystèmes. Afin d’éclairer les déterminants de ce clivage, une enquête à diffusion électronique a été réalisée entre décembre 2015 et mars 2016. Ce document présente l’analyse des réponses et leurs implications.
2 Matériel et méthodes
2.1 Structure de l’enquête et diffusion
Le questionnaire original affichait le double objectif de sonder la communauté scientifique de l’écologie trophique, mais aussi de recenser les forces humaines et les plateformes analytiques en France. Il comportait donc 23 questions au total, dont 15 visaient plus spécifiquement la compréhension du paysage national de l’écologie trophique.
Les six premières questions, à choix multiples, définissaient l’appartenance au champ de l’écologie trophique des personnes interrogées et testaient leur compréhension globale de la notion d’écologie trophique. Il était ainsi demandé à quel niveau l’individu intègre ses recherches dans le cadre de l’écologie trophique (cinq niveaux allant de « pas du tout » à « totalement »), puis la personne sondée était invitée à réévaluer sa réponse après la prise en compte de la définition de l’écologie trophique : « L’écologie trophique se définit comme l’étude du déterminisme et des conséquences écologiques des relations alimentaires et de l’acquisition des ressources nutritives par les êtres vivants. » À la suite de cette première série de questions, les individus hors du champ de l’écologie trophique étaient guidés vers la sortie du questionnaire.
Les cinq questions suivantes, toujours à choix multiples, visaient à caractériser les concepts scientifiques inhérents aux questions de recherche en écologie trophique, les écosystèmes d’étude (c’est-à-dire, les catégories générales subdivisées en écosystèmes plus spécifiques), le type de climat d’étude, les principaux organismes-cibles et les types d’approches scientifiques (par exemple, expérimentales, terrain, in silico). Le nombre de choix était limité à 2 ou 3 par question, mais toutes les questions comportaient, soit un choix libre (en cas d’inadéquation des propositions), soit un choix stipulant l’absence d’objet privilégié. La question 12, à deux niveaux imbriqués de choix (comme pour celle sur les écosystèmes étudiés), renseignait les principaux outils méthodologiques. La dernière question à choix multiples concernait l’identification des verrous ponctuels dans la progression des recherches en écologie trophique.
La dernière série de questions, à champ libre, définissait le niveau hiérarchique, l’identité (non obligatoire) et les coordonnées des personnes sondées, et faisaient place aux commentaires libres. Une courte description de la nature des recherches pouvait aussi être fournie à ce niveau du questionnaire. Le questionnaire a été publié par l’outil d’enquête Inra (LimeSurvey®) et une invitation à participation a été diffusée par messagerie électronique, aux réseaux de contacts, aux laboratoires recensés en écologie, et aux listes de diffusion de sociétés savantes en écologie (Association française de limnologie, réseau GRET, Société française d’écologie…) en décembre 2015. La date limite de participation a été fixée au 31 mars 2016.
2.2 Traitement des résultats
Les réponses aux questions à choix multiples ont été visualisées et analysées au travers d’histogrammes de distribution des réponses, complétés au besoin par un test de chi-deux afin de tester l’homogénéité (hypothèse nulle = distribution homogène ou pas de différences de représentation par groupes) des réponses par catégories. La multiplicité des choix possibles ne permettait pas de réaliser de simples distributions croisées pour évaluer l’existence d’associations privilégiées entre deux catégories (écosystèmes × méthodes, par exemple). Les croisements de QCM ont donc été réalisés par des tableaux de corrélations croisées dont les probabilités sous hypothèse nulle (pas d’associations entre catégories) sont évaluées, avec corrections de Bonferroni au risque α = 0,05 dans le cas de comparaisons multiples.
3 Résultats
3.1 Public sondé
Trois cent trente-six individus ont répondu au questionnaire. Parmi les sondés, 58 % ont spécifié leur laboratoire d’origine, et l’ensemble constitue un panel représentatif des établissements publics à caractère scientifique et technique et des établissements publics à caractère industriel et commercial de la recherche française en écologie et environnement (Universités, UMR INRA et CNRS, IRSTEA, IFREMER et IRD). Parmi les sondés, 75 % n’appartiennent pas au GDR GRET, attestant ainsi la diffusion du questionnaire au-delà des limites du réseau lui-même. Soixante pour cents des enquêtés sont des étudiants de niveau mastère 1 et 2, et 30 % des chercheurs et enseignants-chercheurs titulaires.
3.2 Du concept d’écologie trophique et de son appartenance
Parmi les personnes sondées, 10 % considèrent que leurs recherches n’ont aucune relation avec le champ de l’écologie trophique, tandis que 82 % jugent qu’il s’agit d’un champ qui les concerne au moins en partie (Fig. 1a). Parmi les individus, 75 % révisent leur degré d’implication de l’écologie trophique dans leur recherche après mention de la définition proposée (Fig. 1a, b), attestant ainsi un certain flou du concept même de l’écologie trophique, sa portée, ses tenants et ses aboutissants. Si la distribution générale des réponses ne change que peu après définition (χ2 = 20, p = 0,22), il s’agit essentiellement d’une modération des degrés d’implication, et/ou d’un recentrage des avis initialement extrêmes.

a. Distribution initiale des degrés d’implication de l’écologie trophique dans les recherche des sondés avant (gris foncé) et après (gris clair) explicitation de la définition. b. Modifications relatives des effectifs des différentes catégories après définition. c. Champs auxquels les sondés appliquent des aspects trophiques.
Dans la suite de l’analyse, ne sont conservés que les individus qui considèrent que l’écologie trophique appartient « complètement » ou « en partie » à leurs recherches (301 sondés). À ce stade, nous faisons l’hypothèse que cet échantillon est représentatif de la population des chercheurs en écologie trophique en France.
3.3 Le profil du chercheur en écologie trophique
Trente pour cent de l’échantillon sont des femmes, ce qui est similaire à la proportion observée en 2013 à l’échelle de la recherche nationale [4]. L’écologie trophique concerne, dans la majorité des cas (60 %), une part élevée mais pas unique des travaux de recherche (Fig. 1a). Il s’agit d’une thématique centrale mais pas nécessairement d’une problématique de recherche en soi. La considération des aspects trophiques n’est pas répartie de façon homogène entre différents questionnements scientifiques (χ2 = 114, p < 10−16, Fig. 1c). Ils sont en majorité (30 %) considérés pour leur implication en dynamique des populations et des communautés (niche alimentaire, taux de survie, reproduction, mortalité, compétition inter- et intraspécifique) et pour leurs implications biogéochimiques (20 %, processus de transfert et transformation de la matière, transfert des contaminants, liens avec les fonctions et les cycles). Leurs relations avec le comportement (optimal foraging strategies…), leurs implications évolutives (traits d’histoire de vie et fitness, processus de sélection, niche, régulateur de diversité), leurs applications en bio-indication et biologie de la conservation et les relations trophiques comme un type d’interaction entre espèces/individus (théorie des réseaux d’interactions, topologie…) concernent chacun environ 10 % des recherches. Parmi les sondés, 62 % placent leur recherche sur plusieurs de ces questionnements. Une analyse des corrélations révèle deux principaux pôles d’associations thématiques : comportement–évolution–dynamique et dynamique–interactions. La biogéochimie et la bio-indication/conservation au contraire ne semblent pas présenter d’association claire avec les autres questionnements (figure non montrée).
Soixante huit pour cent des scientifiques considèrent avoir un écosystème d’étude privilégié, attestant que cette recherche est ancrée à un terrain d’étude (Fig. 2a). Les recherches en écologie trophique sont assez équitablement représentées entre les trois grands types d’écosystèmes. En revanche, les chercheurs dont les recherches couvrent au moins deux de ces écosystèmes, classés comme « mixte », restent rares (8 %). Seuls 10 % n’ont pas de type écosystémique privilégié (« aucun/indépendant »), soit parce qu’ils travaillent sur des systèmes théoriques ou de laboratoire (par exemple, modèle drosophile/parasite), ou parce que l’écosystème change selon l’opportunité des questions et/ou données disponibles. Au sein de chaque grand type d’écosystème, il existe des milieux bénéficiant d’un effort plus important de recherche (Fig. 2b–d). Dans les systèmes marins, la zone intertidale est celle qui bénéficie du moins d’attention. En eaux continentales, les eaux souterraines restent extrêmement peu étudiées. En écosystèmes terrestres, les prairies, toundras, forêts et milieux agricoles représentent l’essentiel de l’attention.

a. Écosystèmes-types auxquels appartiennent les milieux d’études privilégiés. b–d. Distribution des milieux d’études au sein des grands types écosystémiques.
Les zones climatiques privilégiées de recherche sont très largement les zones tempérées (54 %) et les climats chauds (méditerranéen, sahélien, arides : 15 %) et tropicaux à subtropicaux (18 %). Les climats froids (boréaux, haute altitude et latitude) sont très largement sous-représentés.
Parmi les organismes-cibles des travaux de recherche, les animaux font l’objet de plus de 60 % des questionnements de recherche (Fig. 3) avec, là encore, une surreprésentation de certains groupes taxonomiques, comme les vertébrés (poissons, mammifères et oiseaux), arthropodes et mollusques.
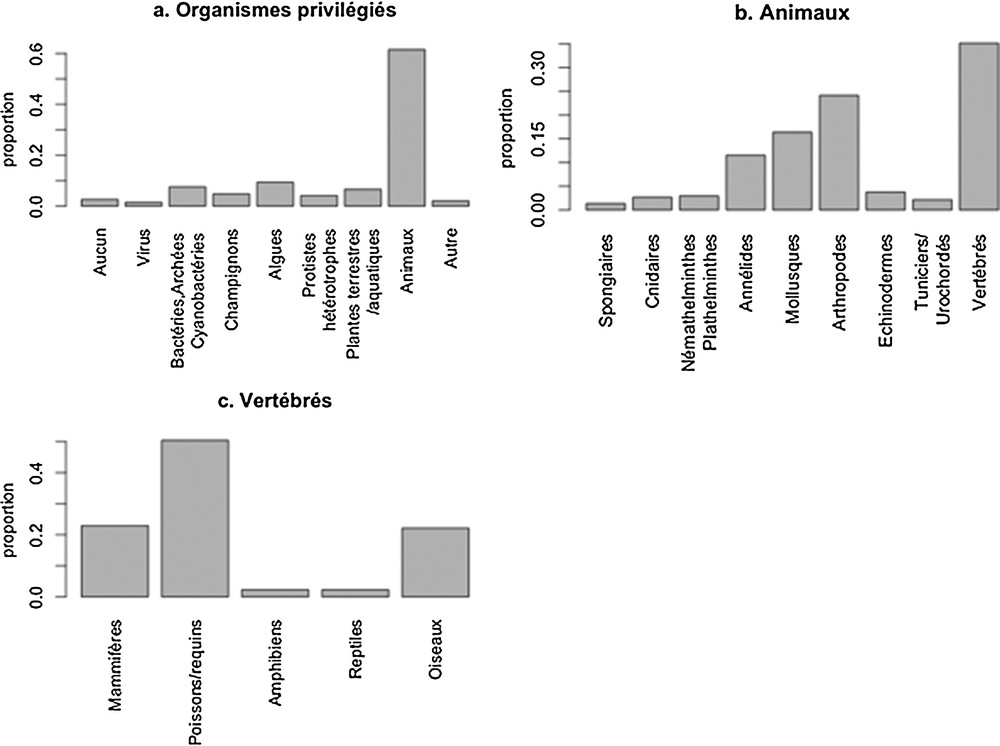
Répartition des organismes-cibles.
Les interactions trophiques font intervenir au moins deux organismes, qui appartiennent ou non aux mêmes catégories taxonomiques. L’analyse des corrélations montre qu’il n’existe pas d’association privilégiée entre différents taxons métazoaires (les animaux). En revanche, les interactions microbiennes constituent un sujet d’étude qui semble relativement déconnecté du compartiment métazoaire (Fig. 4.).
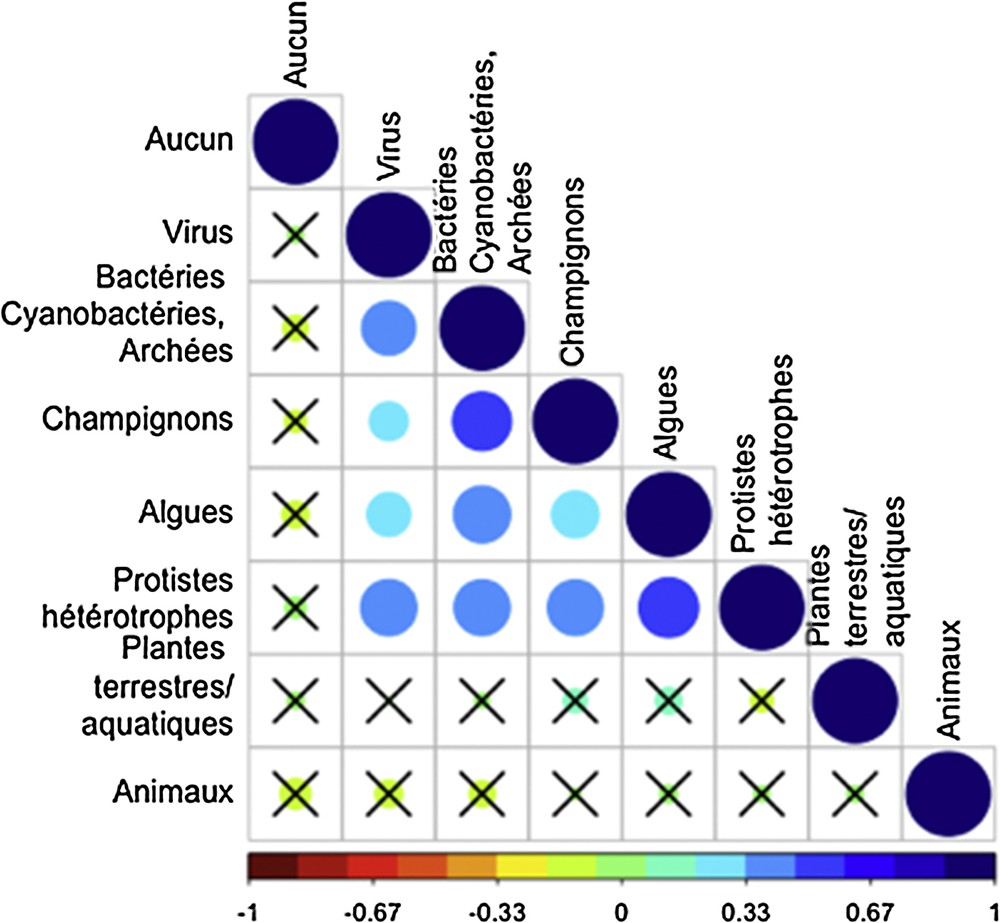
Matrice des corrélations des différents types d’organismes d’étude. La taille et la couleur des cercles est à relier avec la valeur du coefficient de corrélation. Les × indiquent les corrélations non significatives au seuil de 0,05, c’est-à-dire corrigées par les critères de Bonferroni.
La recherche en écologie trophique reste un domaine de terrain (48 %) et d’expérimentation à différentes échelles (33 %), tandis que la modélisation, quel qu’en soit le type, est une approche non négligeable, mais encore minoritaire (20 %, Fig. 5).

Distribution des types d’approches.
Les méthodes appliquées en écologie trophique recouvrent six grandes catégories, assez équitablement représentées, excepté pour les OMICS (par exemple, génomique, protéomique…), dont la sous-représentation reflète probablement la relative nouveauté (Fig. 6).

Distribution des grandes catégories méthodologiques.
Ces méthodes d’analyse des réseaux trophiques sont souvent combinées. L’analyse des contenus stomacaux semble une méthode de base, incontournable quelles que soient l’étude et la méthodologie. D’autres associations privilégiées apparaissent clairement (contenus stomacaux et observations directes du processus alimentaire ; traceurs de biomasse et paramètres écophysiologiques ; modélisations inférentielle et théorique). Les OMICS, en revanche, ne semblent associées à aucune autre méthode en particulier (Fig. 7), et l’analyse fine des résultats individuels confirme cette observation.
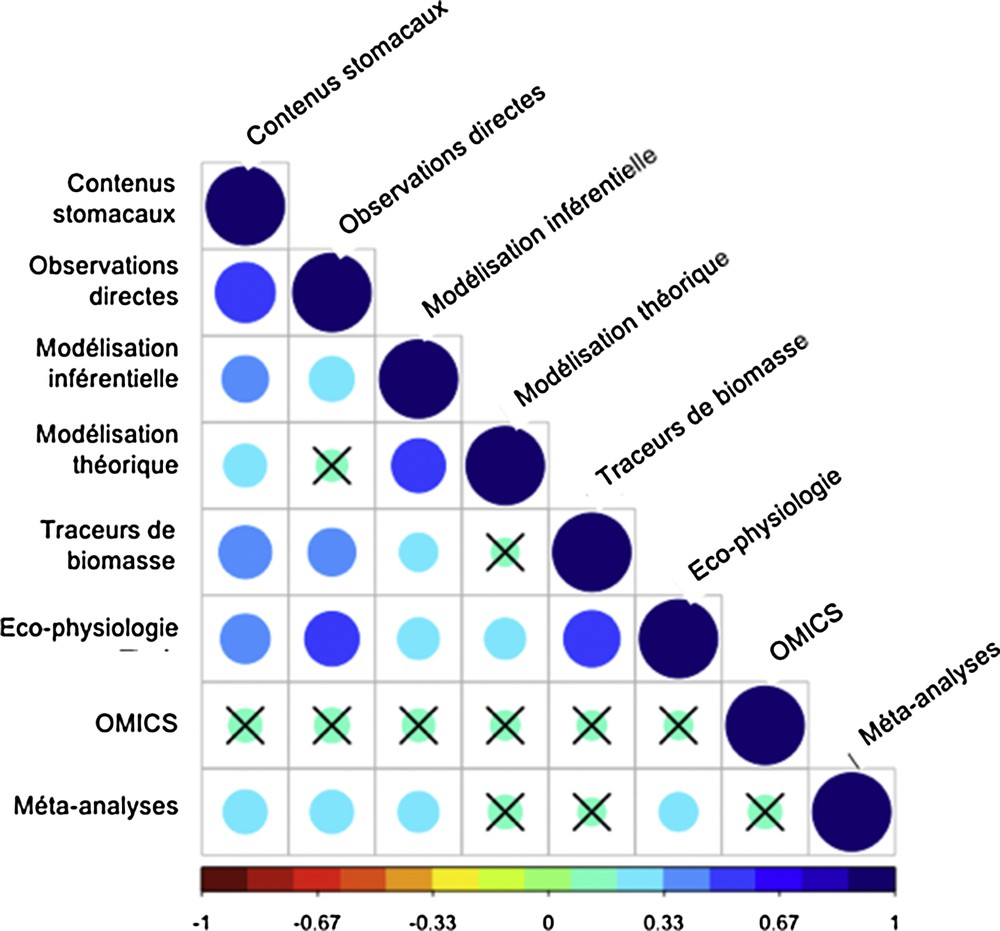
Matrice des corrélations des méthodes. La taille et la couleur des cercles est à relier avec la valeur du coefficient de corrélation. Les croix indiquent les corrélations non significatives au seuil de 0,05, corrigé par les critères de Bonferroni.
3.4 Détection des associations privilégiées entre écosystèmes, méthodologies, organismes et questionnements scientifiques
La suite de l’analyse vise à évaluer s’il existe une ségrégation des communautés de recherche en écologie trophique entre les différents biomes ou organismes modèles, et d’en souligner les possibles causes (enjeux différents sur les différents biomes, ou un certain héritage culturel et/ou méthodologique).
L’analyse des corrélations croisées documente la distribution non aléatoire des questionnements entre les différents écosystèmes d’étude (Fig. 8a–c), confirmant que les questions de dynamique du réseau concernent plus spécialement les écosystèmes aquatiques marins ou dulçaquicoles, tandis que les études des biomes terrestres s’approprient plus souvent les questions d’évolution. La biogéochimie semble un questionnement spécifique aux études qui couvrent plusieurs écosystèmes mixtes (par exemple, connexion milieux terrestres et eaux continentales). Les questionnements relatifs à l’éthologie/comportement alimentaire et aux interactions sont traités sans affiliation à un écosystème type (domaine a-écosystémique). À l’inverse, les questionnements relatifs à la conservation ou à la bio-indication ne sont pas associés de façon privilégiée à une catégorie en particulier. Il s’agit du seul champ disciplinaire de l’écologie trophique qui soit couvert de façon transécosystémique.

Matrice des corrélations, ainsi que de leur significativité, entre (a) questions et écosystèmes, (b) questions et méthodes, ainsi que (c) écosystèmes et méthodes. La taille et la couleur (même échelle que sur la Fig. 7) des cercles est à relier à la valeur du coefficient de corrélation. Les croix × indiquent les corrélations non significatives au seuil de 0,05, corrigé par les critères de Bonferroni.
Il existe aussi une association assez claire et logique entre les questionnements scientifiques et les méthodes. Les questionnements relatifs à la dynamique des populations sont plutôt abordés par des méthodes d’observations directes, combinées à des mesures de paramètres écophysiologiques (fécondité, mortalité, survie…) et des modèles inférentiels. Les questionnements relatifs à la biogéochimie font appel aux méthodes de traçages, naturels ou artificiels, tandis que les études des interactions sont clairement associées à la modélisation inférentielle. Les méthodes basées sur les OMICS, et dans une moindre mesure, la modélisation théorique et les méta-analyses, sont beaucoup moins associées à des questionnements spécifiques.
En dépit de cette nette différenciation des thématiques par type d’écosystème, et d’une logique d’adaptation globale des méthodes (hormis les OMICS) aux questions de recherche, il n’existe que peu d’associations privilégiées entre méthodes et écosystèmes (Fig. 8b). Les recherches en milieu marin semblent privilégier les méthodes par traçages, l’examen des contenus stomacaux et l’écophysiologie, reflétant peut-être un héritage culturel et/ou la faisabilité et la facilité de mise en œuvre de ces méthodes au sein de ces écosystèmes ou sur les organismes concernés. Les méthodes de modélisation s’appliquent indifféremment aux différents écosystèmes ou à des systèmes théoriques, et montrent donc une association à « aucun écosystème » et mettent en lumière la transversalité de ces méthodes dans la communauté scientifique. Les communautés de chercheurs en écologie trophique partagent assez bien un bagage similaire d’expertise méthodologique.
Le choix des organismes-cibles dépend peu des questionnements scientifiques (Fig. 9a), hormis pour les animaux dont la compréhension de la dynamique reste une question clé, et du rôle du compartiment microbien dans les processus biogéochimiques. Ainsi, les classes d’organismes-cibles varient peu entre biomes d’étude, hormis une sous-représentation naturelle des études sur les algues en milieu terrestre (Fig. 9b). Reste néanmoins le fait que les études trophiques relatives au compartiment microbien sont assez exclusives aux milieux dulçaquicoles, tandis que les études sur les animaux sont sur-représentées en milieu marin.
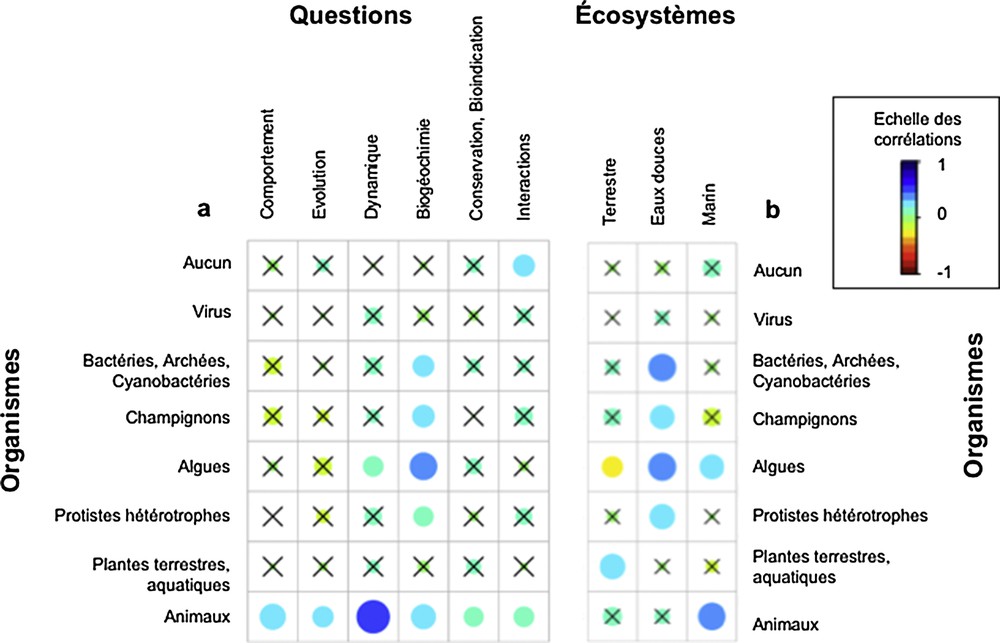
Représentation des corrélations entre organismes et (a) questionnements ou (b) écosystèmes d’étude.
Ce constat est confirmé par le calcul du niveau trophique moyen des espèces étudiées, pour chaque type d’écosystèmes d’étude (Fig. 10). Ce niveau trophique moyen est très similaire pour les écosystèmes terrestres, dulçaquicoles et mixtes, et se place à une valeur de 1,5, typique d’une attention moyenne autour des interactions producteur–consommateur primaires (« plantes-herbivores »). Si le niveau moyen d’intérêt pour ces écosystèmes se place assez bas dans la chaîne alimentaire, la distribution autour de cette valeur moyenne démontre néanmoins une large couverture du spectre trophique. En moyenne, les chercheurs issus des écosystèmes marins placent leur attention à un échelon trophique supérieur par rapport à ceux attachés à d’autres catégories d’écosystèmes, au niveau des interactions entre consommateurs primaires et secondaires (« consommateurs herbivores/détritivores-prédateurs »).
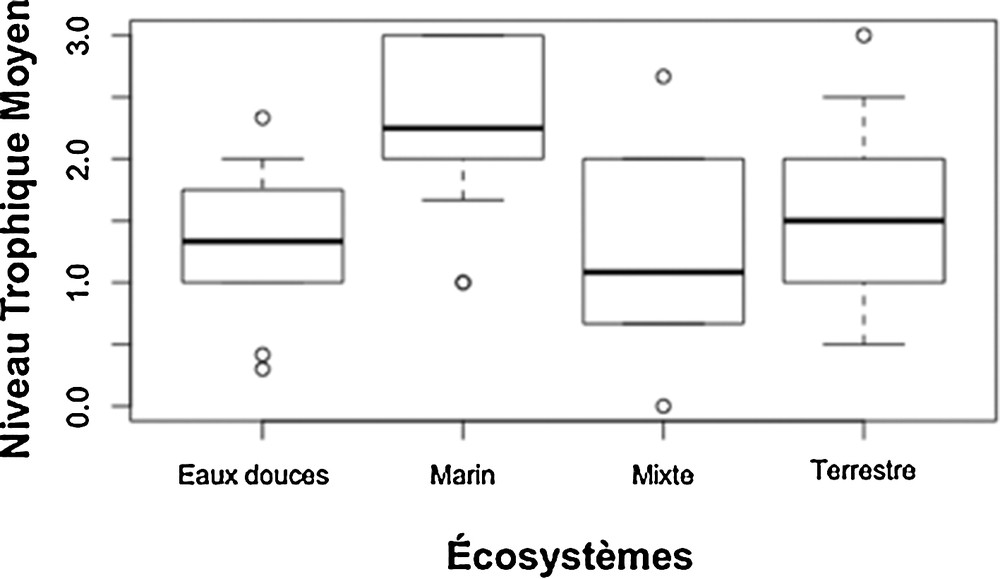
Distribution des niveaux trophiques moyens d’étude par grandes catégories d’écosystèmes.
Il existe aussi des associations nettes entre questionnements et type d’approches privilégiées (Fig. 11). Le terrain reste une base à tout type de questionnements, hormis l’étude des interactions, mais la biogéochimie est traitée spécifiquement par des études expérimentales ainsi que du terrain, avec peu d’associations avec des méthodes de modélisation. Il ne semble pas y avoir de question privilégiée dans les approches de modélisation (tant théorique qu’inférentielle), qui sont peu ou pas corrélées aux différentes classes de questions. Il résulte des associations détectées entre questions et approches, une ségrégation là encore entre biomes. Les approches expérimentales semblent davantage le fait des communautés de chercheurs du domaine des milieux terrestres ou dulçaquicoles, tandis que ces approches sont sous-représentées dans celui des milieux marins. Les observations de terrain semblent plus représentées dans les milieux aquatiques que dans les milieux terrestres.

Matrices des corrélations entre type d’approches (micro-méso-mégacosmes/modélisations inférentielles et théoriques, terrain) et a) type de questionnements et b) biomes d’étude.
3.5 Les verrous identifiés dans la communauté
Sans surprise, les participants identifient le manque de salaires/personnel comme principal obstacle à leur progression scientifique en France : 60 % évoquent un manque de moyens humains, plutôt que techniques ou d’équipement (Fig. 12). Le problème qui relève d’une communauté en écologie trophique mal identifiée, noté comme important au regard de l’INEE, apparaît ainsi assez négligeable face au manque de financement en ressources humaines auprès des participants.
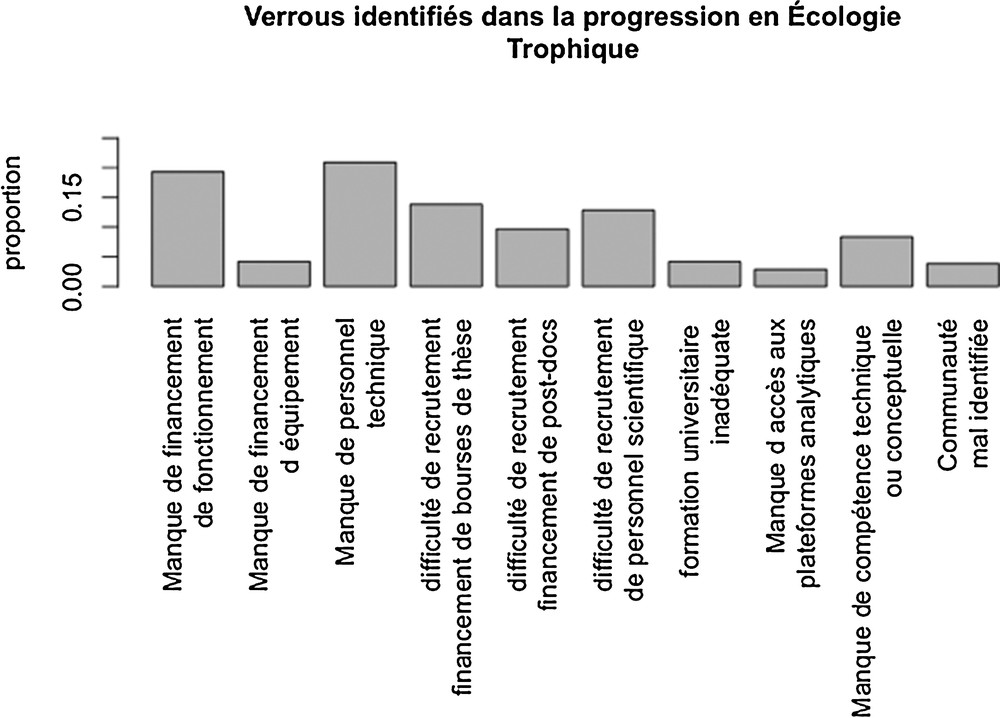
Verrous de progression identifiés.
4 Discussion
4.1 Une bonne identification à la thématique de l’écologie trophique
« Montre-moi ce que tu manges et je te dirai qui tu es. Pareille question devrait s’adresser au préalable à tout insecte dont on étudie les mœurs, car du plus gros au moindre dans la série animale, le ventre est le souverain du Monde ; les données fournies par le manger dominent les autres documents de la vie [5] ».
Dans le cadre d’une enquête menée parmi 388 scientifiques de la British Ecological Society, Sutherland et al. [6] ont identifié les 100 questions considérées d’importance fondamentale en écologie générale. La question trophique transparaît dans nombre des questionnements soulevés dans cette réflexion [6] (c’est-à-dire évolution, populations, communautés et écosystèmes). L’écologie trophique demeure une thématique centrale à laquelle se réfèrent la majorité des questionnements en écologie, mais également certaines questions sociétales majeures comme la dynamique future quantitative et qualitative de nos ressources alimentaires dans un contexte de changement global. Le sondage ici présenté a été diffusé de sorte à atteindre l’ensemble de la communauté des chercheurs français en écologie. Il est néanmoins fort probable que les individus ayant pris le temps de le remplir aient été ceux qui se sentaient d’emblée concernés par les thématiques de l’écologie trophique. Le pivot avéré que constitue la modélisation dans l’étude des processus biogéochimiques (marins comme terrestres) n’apparaît pas dans la communauté des sondés. Cette sous-représentation soulève la question d’une possible fragmentation de la communauté des biogéochimistes entre les unités sous tutelle INEE et celles sous tutelle INSU qui, de fait, n’auraient pas été touchées par notre requête. Dans ce sens, cette étude ne saurait fournir un panorama de la place de l’écologie trophique dans la recherche en écologie, à l’échelle nationale. Émerge toutefois un fort sentiment d’appartenance à cette thématique, qui transcende de nombreuses questions écologiques au sens plus général. Les résultats de l’analyse des réponses confirment les clivages supposés préalablement de façon plus empirique et permettent d’en dessiner les causes ainsi que d’identifier des opportunités pour construire des ponts.
4.2 Il y a peu de recouvrement entre écologie trophique et écologie microbienne
Un des constats les plus flagrants concerne la rareté des écologues microbiens qui associent leurs recherches à la thématique de l’écologie trophique. Ainsi, l’écologie trophique serait vue comme celle des interactions macroscopiques plantes–animaux. Seuls 20 % de sondés mentionnent les microbes (protistes, algues, bactéries, archées champignons) parmi leurs organismes d’étude. L’analyse croisée confirme que les associations microbiennes sont étudiées indépendamment de celles avec les organismes pluricellulaires. Les résultats suggèrent aussi que ces microbiologistes, qui intègrent le champ thématique de l’écologie trophique, s’attachent à des questions de biogéochimie avec une surreprésentation des milieux aquatiques, bien que ces associations restent à prendre avec retenue du fait de la faible taille de la population des microbiologistes dans le public sondé. À titre de comparaison, les modélisateurs, qui représentent aussi 20 % des sondés qui s’identifient au champ de l’écologie trophique, couvrent un grand nombre des questionnements, des biomes et des organismes. Autrement dit, si les modélisateurs sont encore minoritaires, leurs travaux ne s’exercent pas indépendamment des activités des chercheurs non modélisateurs en écologie trophique.
Les microbes contiennent la majorité de la diversité génétique planétaire, la biomasse microbienne planétaire est du même ordre de grandeur que celles de l’ensemble des plantes, tandis que le stockage microbien des nutriments N et P est 10 fois supérieur à celui des plantes [7]. Par leur métabolisme, ils sont des acteurs majeurs des grands cycles biogéochimiques. Les relations des microbes vis-à-vis de leur source nutritionnelle, de leurs prédateurs, de leurs hôtes ou de leurs symbiontes sont autant de mécanismes trophiques importants régulant leur dynamique, diversité et évolution [7]. L’absence de zones de contact entre les communautés de l’écologie microbienne et trophique limite donc forcément notre capacité à aborder ces processus dans leur globalité, alors qu’ils sous-tendent des enjeux sociétaux majeurs (le rôle de la biodiversité et les liens aux services écosystémiques, la nécessité de contrôler les flux en nutriments, favoriser les stockages de carbone).
Ce constat à l’échelle française s’observe aussi empiriquement au niveau de la recherche internationale [6]. Alors que la fin des années 1980 avait été marquée par l’avènement du concept de boucle microbienne, Azam, Fenchel et al. [8], Sherr and Sherr [9,10], Pomeroy, Williams et al. [11] ont tous prôné l’intégration à part entière des acteurs microbiens non photosynthétiques dans la conception des réseaux trophiques, au moins dans les milieux marins et lacustres. Dans les sols, les processus microbiens de recyclage de la matière organique pourraient être de même ampleur que dans les eaux douces ou marines [12]. Pourtant, 30 ans après la conceptualisation de la boucle microbienne, les microbes semblent avoir bel et bien disparu des études trophiques. La volonté du GRET est de remettre en lumière, par des communications de synthèse, le rôle des microbes en écologie trophique, de révéler la dichotomie des conceptions entre écosystèmes, afin de souligner l’importance cruciale du rapatriement des microbes dans les concepts trophiques.
Des raisons de cette ségrégation entre microbienne et écologie trophique peuvent être envisagées, au regard de réflexions récentes et de l’analyse présentée, ainsi que de la littérature internationale. Une première raison pourrait être conceptuelle : de nombreux concepts propres à l’écologie trophique (relation prédateur–proie, sélection, niveau trophique) repose sur la notion d’espèces, concept caduque lorsqu’on tente de l’étendre au monde microbien [13]. Les tests des théories macro-écologiques au monde microbien restent toutefois une question fondamentale identifiée par Sutherland et al. [6]. L’isolement de ces champs de recherche pourrait aussi en partie découler d’une certaine ségrégation des méthodes. L’écologie microbienne a été très récemment révolutionnée par l’avènement et le développement exponentiel des méthodes de séquençage et de bio-informatique à haut débit. Si la démocratisation de ces méthodes a permis de véritablement ouvrir la boîte de Pandore de la diversité microbienne, jusque-là largement sous-estimée, elle a aussi conduit à détourner les microbiologistes des concepts de l’écologie même [14]. Pendant une décennie, l’attention des microbiologistes a été centrée sur le décryptage de cette biodiversité encore inconnue, les développements méthodologiques associés aux méthodes moléculaires et au traitement de l’information générées en haut débit, les éloignant de fait du reste des autres communautés d’écologues [14]. Dans un sens, les avancements technologiques, s’ils ont permis aux analyses de devenir plus complexes et plus sophistiquées, ne garantissent pas nécessairement une progression de la compréhension, ni le bouleversement des concepts [15]. L’écologie générale a été qualifiée, par provocation, de « science à la traîne » par rapport à la recherche biomédicale, cette disqualification ayant attribuée à sa tendance naturaliste, qui chercherait à décrire plutôt qu’à comprendre les processus et lois générales en jeu [16]. Comme Swihart et al. [17], nous appuyons sur la nécessité d’identifier les forces en présence avant de pouvoir prétendre à comprendre les processus, et donc légitimons l’étape obligatoire, mais pas suffisante, de décryptage de la biodiversité. Le développement méthodologique atteint maintenant un relatif plateau, devant permettre relatif retour de la microbiologie vers des concepts plus généraux. On peut donc à présent espérer que les différentes communautés puissent à nouveau se rejoindre.
4.3 La recherche en écologie trophique est territoriale
Au sein même de l’écologie trophique, il existe une ségrégation de la communauté essentiellement dictée par l’écosystème d’étude. Sept sondés sur dix peuvent s’identifier à un type d’écosystème unique d’étude, et on observe une territorialité assez claire : les limnologues basés au pied des Alpes travaillent sur les lacs alpins, les écologues de Banyuls travaillent en Méditerranée, ceux de Brest sur la rade, ceux de la Rochelle sur les zones intertidales… Les climats et écosystèmes d’étude privilégiés sont ceux traditionnels de France métropolitaine (prairies, forêts, paysages agricoles, rivières, lacs, plateau océanique continental) ou d’outre-mer. Cette territorialité des recherches en écologie trophique, ainsi que la proximité des zones d’étude, rappellent que la recherche en écologie trophique est d’abord une recherche de terrain. La régionalisation des recherches en écologie trophique découle en partie d’enjeux socio-économiques différents portés par les biomes étudiés face aux changements globaux. Les questions de dynamique des populations en milieu marin se rattachent aux enjeux du maintien des pêcheries [18]. La biogéochimie est un thème important dans les eaux continentales, dont la qualité dépend des transferts de nutriments dans les bassins versants [19]. Une partie du clivage géographique observé est donc légitime, puisque l’écologie trophique en France porte des applications assez directes à la gestion et à la préservation de l’environnement. À ce titre, la recherche française en écologie trophique se distingue de la recherche internationale en écologie générale qui n’est, quant à elle, que peu dirigée par des problèmes environnementaux et sociétaux [15]. La territorialité de la recherche en écologie trophique en France serait ainsi en partie le reflet de son implantation importante dans des établissements à vocation de recherche appliquée (Inra, Ifremer, Irstea, Cirad, IRD…).
Ces enjeux sociétaux ne sont toutefois pas le seul facteur de structuration des communautés en écologie trophique. Par exemple, la qualité des eaux des zones côtières représente un enjeu sociétal majeur, dont la compréhension demande de s’attarder sur les plus bas niveaux des réseaux trophiques (producteurs primaires = niveau 1). Pourtant, le niveau trophique moyen étudié dans les milieux marins est bien plus proche des prédateurs supérieurs, indépendamment des différents enjeux. Par ailleurs, la recherche sur les questions d’évolution est de nature plus fondamentale et, de ce fait, peu dépendante des enjeux sociétaux. Elle fait pourtant quasi-exclusivement l’objet d’études de biomes terrestres, et atteste un partage inégal des concepts de l’écologie théorique entre les différents biomes d’étude. La régionalisation des recherches en écologie fondamentale semble donc le fait d’un héritage culturel que des instruments de réseautage tels que le GRET, qui dépassent le maillage géographique initial des unités de recherche, peuvent permettre d’homogénéiser. Les résultats montrent, par ailleurs, que, dans l’ensemble, les écologues trophiques ont un bagage méthodologique assez commun, quel que soit leur écosystème d’étude, et la modélisation, notamment théorique, est affranchie de la nature de l’écosystème. La diversité des méthodes en écologie trophique ne sera donc a priori pas un obstacle au rassemblement des écologues trophiques à terrain préférentiel (terrestres, marins, dulcicoles), non spécifiquement affiliés à un écosystème ou au contraire aux approches transécosystémiques. Un des leviers d’action d’un GDR tel que le GRET pourrait être alors de promouvoir des études transversales entre les différents biomes visant à tester les concepts de l’écologie fondamentale (par exemple, le rôle de la diversité verticale sur la relation biodiversité fonction, le rôle de la qualité nutritionnelle sur les transferts trophiques, la question du priming effect…). Effectivement, les tests expérimentaux à grande échelle des théories reliant le rôle de la biodiversité aux fonctions et services écosystémiques ont été essentiellement le fait des milieux prairiaux (programme BIODEPTH [20]), sans démarche équivalente à ce jour sur les milieux d’eaux continentales ou marines, en dépit d’enjeux tout aussi importants. Les nouvelles infrastructures expérimentales à grande échelle de l’INEE ou des investissements d’avenir (Ecotron, Planaqua, les stations d’écologie expérimentale) et les observatoires en environnement (zones ateliers, observatoires-hommes milieux, SOERE, OSU, etc.) de l’INEE ou d’autres instituts (Insu, CNRS, Inra) et Allenvi sont des outils idéaux pour mener en parallèle des tests expérimentaux sur de multiples écosystèmes, et donc d’évaluer la pertinence des concepts de façon transécosystémique. Reste néanmoins que le financement est identifié comme le verrou majeur quant à l’avancement des travaux en écologie trophique, avec un accent des sondés sur le manque de personnel technique et d’étudiants. Ainsi, si l’outil existe, son utilisation resterait limitée par le manque d’obtention de financements, et plus particulièrement de ceux permettant de recruter les moyens humains nécessaires au fonctionnement de ces infrastructures dans la durée.
4.4 La recherche en écologie trophique est limitée aux frontières de l’écosystème
Les notions d’« écosystèmes » et de « communautés » sont unanimement considérées comme des concepts fondateurs et prioritaires de la recherche écologique (rapporté dans Carmel et al., 2013 [15]). Pourtant, l’écologie générale reste quantitativement cantonnée à une écologie des populations, avec > 60 % des études qui ne concernent qu’une seule espèce [15]. L’écologie trophique concerne, par essence, les interactions entre les espèces, reliant ainsi les questions de populations aux communautés, voire aux écosystèmes via les réseaux trophiques et l’écologie fonctionnelle. La communauté des scientifiques français(es) en écologie trophique est large et diversifiée. Si elle est organisée et connectée, elle constitue un vivier qui a le potentiel de diriger et de stimuler l’évolution de l’écologie générale vers l’échelle de l’écosystème. S’y ajoute sa force de modélisation. Alors que la proportion d’articles en écologie basée sur de la modélisation stagne à 12 % depuis 30 ans, la communauté des chercheurs en écologie trophique en France contient plus de 20 % de modélisateurs, dont les préoccupations couvrent l’ensemble des questionnements abordés dans l’enquête. Les approches modélisatrices sont au cœur, non seulement des changements d’échelle (passage d’échelles spatiales locales à régionales), mais aussi des tests d’hypothèses (passage du « quoi, qui ? » naturaliste au « pourquoi, comment ? » mécaniste [16]). Le GRET a le potentiel et la volonté de démocratiser l’usage et la collaboration en modélisation, créant ainsi un courant inspirant pour la recherche nationale et internationale.
Il faut enfin souligner la faiblesse des recherches aux interfaces entre écosystèmes, les approches mixtes combinant deux types de biomes représentant moins de 10 % des réponses, et les recherches sur les écotones (milieu hyporhéique, zones intertidales) moins de 5 % des recherches. L’écologie trophique, reflétant là encore une tendance plus générale de l’écologie, reste donc encore très empreinte du concept originel d’écosystème, comme unité spatiale délimitée par des frontières conventionnelles [21]. Si le concept d’écosystème a conduit à une indéniable progression de la recherche en écologie, il apparaît maintenant que les nouveaux enjeux sociétaux demandent de le dépasser en s’affranchissant de ses frontières [22,23]. Le rôle des transferts latéraux d’espèces et de la connectivité et diversité de l’habitat sur le maintien de la biodiversité, la vulnérabilité aux espèces invasives, la mobilité inter-écosystèmes des grands prédateurs, les transferts de nutriments et de carbone entre écosystèmes et leur rôle dans le maintien de la production des milieux ou de la qualité de l’eau, sont autant de questions qui exigent une approche méta-écosystémique [24,25]. Là encore, des réseaux comme le GRET peuvent faire germer dans les esprits des membres la nécessité de décloisonner des recherches. Les financements de ces réseaux restent toutefois trop limités pour soutenir directement de nouveaux travaux.
5 Conclusion
Comme pressenti en amont de cette enquête, la communauté des chercheurs en écologie trophique est divisée entre les organismes et les grands biomes d’étude. Une part de ce clivage peut être attribuée à la spécificité de certains enjeux sociétaux. Les résultats révèlent que l’héritage culturel y joue toutefois un rôle crucial et constitue un frein à la progression des connaissances, tant fondamentales qu’appliquées, dans le domaine de l’écologie trophique. Les initiatives de réseautage telles que le GRET constituent donc un levier d’action potentiel sur ce clivage, en le mettant en lumière auprès de cette communauté, en identifiant les forces en présence et en soulignant les bénéfices, l’innovation et la possibilité de dépasser ces clivages, notamment auprès des jeunes chercheurs. En pratique, l’expérience au sein du GRET a montré que des discussions, dont l’origine fut amorcée par le hasard des rencontres entre chercheurs d’horizons différents, ont pu mettre en lumière des liens précis à établir et des espaces à combler dans notre compréhension des processus. S’en remettre au hasard n’apparaît bien évidemment pas intellectuellement satisfaisant, mais mettre celui-ci de côté serait une erreur. Aussi, le GDR GRET cherche à favoriser au maximum les interactions entre chercheurs et à donner du temps aux discussions durant ses journées annuelles.
En ce sens, le GDR GRET vise à créer un espace de « Slow Science », hors de la logique productiviste de la publication spécialisée qui constitue le critère de ladite « excellence » [25,26]. Notre communauté prend ici le temps de se poser les questions fondamentales autrement laissées en suspens, notamment parce qu’elles ne pourraient faire l’objet d’une récompense immédiate sous forme de publications [26,27]. Dans cet objectif de créer du sens à cette recherche en écologie trophique, les présentations volontairement très courtes des travaux de recherche des participants ne constituent qu’un prétexte pour amorcer les discussions d’où pourront découler, dans le cadre d’une vision transécosystémique, l’identification et la mise en place d’ateliers thématiques spécifiques centrés autour d’une problématique, d’un concept, d’une barrière méthodologique voire d’un groupe taxonomique ou fonctionnel, etc. Ces ateliers thématiques rassemblant un groupe plus restreint de participants évoluent par la suite à leur guise en définissant leurs objectifs, tels que la rédaction de papiers d’opinion ou de projets de recherche permettant des expériences communes. À cet égard, d’autres initiatives telles que les infrastructures expérimentales de l’INEE et les observatoires en environnement sont autant d’atouts sur lesquels capitaliser pour reconnecter l’écologie trophique. Transformer l’essai demandera néanmoins, à terme, que des programmes de recherche émergent de ce réseau, en dépit du goulet d’étranglement que constitue le financement de la recherche aujourd’hui en France.
Remerciements
Nous remercions le relecteur anonyme pour son avis éclairé sur ce manuscrit. Les auteurs remercient l’ensemble de leurs collègues, pour avoir accepté d’être, pour une fois, les cobayes de cette grande expérience transécosystémique, ainsi que l’INEE et l’Inra pour les financements accordés au GDR CNRS 3716, sans oublier J. Gret pour les discussions toujours aussi stimulantes que nous avons eues avec lui.
1 http://www.cnrs.fr/inee/, accès le 2/06/2017.