


 CC-BY 4.0
CC-BY 4.0
When the sequence of the Bacillus subtilis genome was completed in 1997, its completeness was questioned by scientific colleagues who found it difficult to accept that the gene encoding the essential and most important ribonuclease in Escherichia coli in terms of numbers of substrates (RNase E), was missing. Indeed, the full genome sequence led to the realisation that not only was the gene encoding RNase E (rne) absent from B. subtilis, but of the ∼15 ribonucleases known in E. coli at that time, only a handful had homologs in B. subtilis (RNase P, RNase III, RNase PH, PNPase, and RNase R). This opened up a whole new field of research for our laboratory, and for others, trying to find the missing components of the B. subtilis RNA maturation and degradation machinery. Today, B. subtilis has been clearly established as an alternative model for RNA metabolism to the E. coli paradigm. In this review, we will describe these two ends of the phylogenetic spectrum and show how two opposing gradients, representing the fading in or fading out of specific ribonucleases, can account for the vast majority of the evolutionary space between these two organisms that separated 1–3 billion years ago.
The amount of any RNA in the cell and thus potentially the amount of protein it encodes is a balance between its rate of synthesis and degradation. While the transcription and translation machinery are remarkably well conserved in bacteria, the RNA degradation machinery is not. In bacteria, RNA is degraded by a combination of endo- and exo-ribonucleases that either cut RNA internally, or remove one nucleotide (nt) at a time from the 5′ or 3′ end. RNase activity can be modulated both at the substrate level (e.g. through changes in its secondary or tertiary structure conformation), and by the presence of certain chemical groups either at the 5′ end (e.g. 5′ triphosphate groups, NAD caps, NpnN caps, CoA caps) or internally (e.g. m6-A, pseudouridine (𝜓) modifications) [1, 2, 3, 4, 5]. It can also be modulated by protein co-factors or by post-translational modification (e.g. acetylation, phosphorylation) [6, 7]. In addition to degrading RNA, RNases can also paradoxically stabilize RNA, by eliminating nucleotides that could potentially recruit degradative enzymes. This is typically the case for the so-called stable RNAs, e.g. ribosomal RNA (rRNA) and transfer RNA (tRNA), that are generally synthesized in the form of precursors that are trimmed at both their 5′ and 3′ ends to yield the final stable and functional molecules [8]. A single RNase can have both stabilizing (maturation/processing) and degradation functions depending on the RNA substrate and whether its product becomes more or less sensitive to other RNases.
There are currently about 40 bacterial RNases known, with the vast majority having been discovered in either E. coli or B. subtilis. E. coli currently has 27 known RNases, while B. subtilis has 21. Most are involved in house-keeping operations such as mRNA decay and stable RNA maturation and their functions are summarized in Table 1. Several belong to the so-called toxin family of RNases. Indeed, the major difference in the total number of RNases between E. coli and B. subtilis is that E. coli has at least ten known toxin RNases [9], while B. subtilis so far only has one [10]. The expression of toxin–antitoxin (TA) pairs is typically induced under different stress conditions and release of the toxin from an inhibitory complex with its antitoxin relies on the rapid degradation of the antitoxin. Once the toxin is released, it acts as an endoribonuclease, cleaving RNA in a sequence specific manner, typically with short recognition sequences of 4–6 nts, that occur very frequently. TA modules are thought to be transferred horizontally and are often associated with mobile elements, some of which have become cryptic over time.
Ribonucleases found in bacteria
Enzyme type | Principle function(s) | |
|---|---|---|
| Core set | ||
| RNase III | Endoribonuclease | Cleavage of double stranded RNA: rRNA precursors, mRNA, antisense RNA |
| RNase H2 | Endoribonuclease | Cleavage of DNA/RNA hybrids |
| YbeY | Endoribonuclease | Processing of 16S rRNA 3′ end |
| RNase P | Endoribonuclease | Processing of tRNA 5′ ends |
| PNPase | 3′-exoribonuclease | Phosphorolytic degradation of mRNA, rRNA. Protection of sRNA |
| RNase R | 3′-exoribonuclease | Hydrolytic degradation of rRNA, mRNA |
| RNase PH | 3′-exoribonuclease | Phosphorolytic processing of tRNA 3′ ends |
| “E. coli” set | ||
| RNase E | Membrane associated endoribonuclease | Cleavage of mRNA, sRNA, processing of tRNA, 16S and 5S rRNA |
| RNase G | Endoribonuclease | Cleavage of mRNA, processing 16S rRNA |
| RNase H1 | Endoribonuclease | Cleavage of DNA/RNA hybrids |
| RNase I | Endoribonuclease | Periplasmic RNase, degradation of rRNA following membrane stress |
| Oligoribonuclease | 3′-exoribonuclease | Degradation of short oligonucleotides, end products of other 3′–5′ exoribonucleotides |
| RNase T | 3′-exoribonuclease | Hydrolytic processing of tRNA 3′ ends |
| RNase D | 3′-exoribonuclease | Hydrolytic processing of tRNA 3′ ends |
| RNase II | 3′-exoribonuclease | Hydrolytic processing of tRNA 3′ ends, shortening of mRNA 3′ ends |
| RNase AM | 5′-exoribonuclease | 5′ processing of 16S, 23S and 5S rRNA |
| “B. subtilis” set | ||
| RNase Y | Membrane associated endoribonuclease | Cleavage of mRNA, processing of RNase P RNA, sRNA |
| RNase Z | Endoribonuclease/3′-exoribonuclease | 3′ processing of tRNA |
| RNase H3 | Endoribonuclease | Cleavage of DNA/RNA hybrids |
| RNase M5 | Endoribonuclease | Processing of 5S rRNA |
| Mini-RNase III | Endoribonuclease | Processing of 23S rRNA, type II self-cleaving ribosymes |
| Rae1 | Endoribonuclease | Cleavage of mRNA through association with ribosome |
| YhcR | Membrane associated endoribonuclease | unknown |
| Bsn | Endoribonuclease | Degradation of extracellular RNA |
| RNase J | 5′–3′ exoribonuclease/ endoribonuclease | Degradation of mRNA, sRNA, processing of 16S rRNA 5′ end |
| NanoRNase A | 3′–5′ exoribonuclease | Degradation of short oligonucleotides, end products of other 3′–5′ exoribonucleotides |
| NanoRNase B | 3′–5′ exoribonuclease | Degradation of short oligonucleotides, end products of other 3′–5′ exoribonucleotides |
| YhaM | 3′–5′ exoribonuclease | 3′ processing of tRNA, shortening of mRNA 3′ ends |
Phylogeny of ribonucleases in bacteria. This is an update and new representation of Ref. [11] showing the first 38 model organisms sequenced, a measure of their scientific importance. (A) The “core set” of RNases conserved in most bacteria. (B) Distribution of the “E. coli set” and “B. subtilis set” of RNases across the phylogenetic tree. (C) Distribution of horizontally transferred toxin RNases across the phylogenetic tree. RNase abbreviations are as follows: III = RNase III; P = RNase P; H2 = RNase HII; H1 = RNase HI; H3 = RNase HIII; E = RNase E; G = RNase G; EG = RNase E/G; YbY = YbeY; I = RNase I; Z = RNase Z; M5 = RNase M5; Y = RNase Y; mIII = Mini-III; Rae = Rae1; Maz = MazF; Rel = RelE; Yoe = YoeB; Hig = HigB; YfQ = YafQ; Yhv = YhaV; LS = RNase LS, Mqs = MqsR; YfO = YafO; Bsn = RNase Bsn; Bar = Barnase; AM = RNase AM; R = RNase R; Pnp = polynucleotide phosphorylase; PH = RNase PH; D = RNase D; Orn = oligoribonuclease; II = RNase II; T = RNase T; J = RNase J; BN = RNase BN; Yha = RNase YhaM. Numbers in superscript refer to the number of orthologs present when identified. Organism abbreviations are as follows: Eco = Escherichia coli; Sty = Salmonella typhimurium; Ype = Yersinia pestis; Vch = Vibrio cholera; Hin = Haemophilus influenza; Xfa = Xylella fastidiosis; Pmu = Pasteurella multocida; Bsp = Buchnera sp.; Pae = Pseudomonas aeruginosa; Nme = Neisseria meningitidis; Rpr = Rickettsia prowazekii; Atu = Agrobacterium tumefaciens; Sme = Sinorhizobium meliloti; Mlo =Mesorhizobium loti; Hpy Helicobacter pylori; Cje =Campylobacter jejuni; Ctr =Chlamydia trachomatis; Cmu = Chlamydia muridarum; Tpa = Treponema pallidum; Bbu = Borrelia burgdorferi; Sco = Streptomyces coelicolor; Mtu = Mycobacterium tuberculosis; Mle = Mycobacterium leprae; Uur = Ureaplasma urealyticum; Mge = Mycoplasma genitalium; Mpe = M. pneumoniae; Cac = Clostridium acetobutylicum; Lla = Lactococcus lactis; Spe = Streptococcus pneumoniae; Spy = Streptococcus pyogenes; Bha = Bacillus halodurans; Lin = Listeria innocua; Sau = Staphylococcus aureus; Bsu = Bacillus subtilis; Ssp = Synechocystis sp.; Dra = Deinococcus radiodurans; Tma = Thermotoga maritima; Aao = Aquifex aeolicus.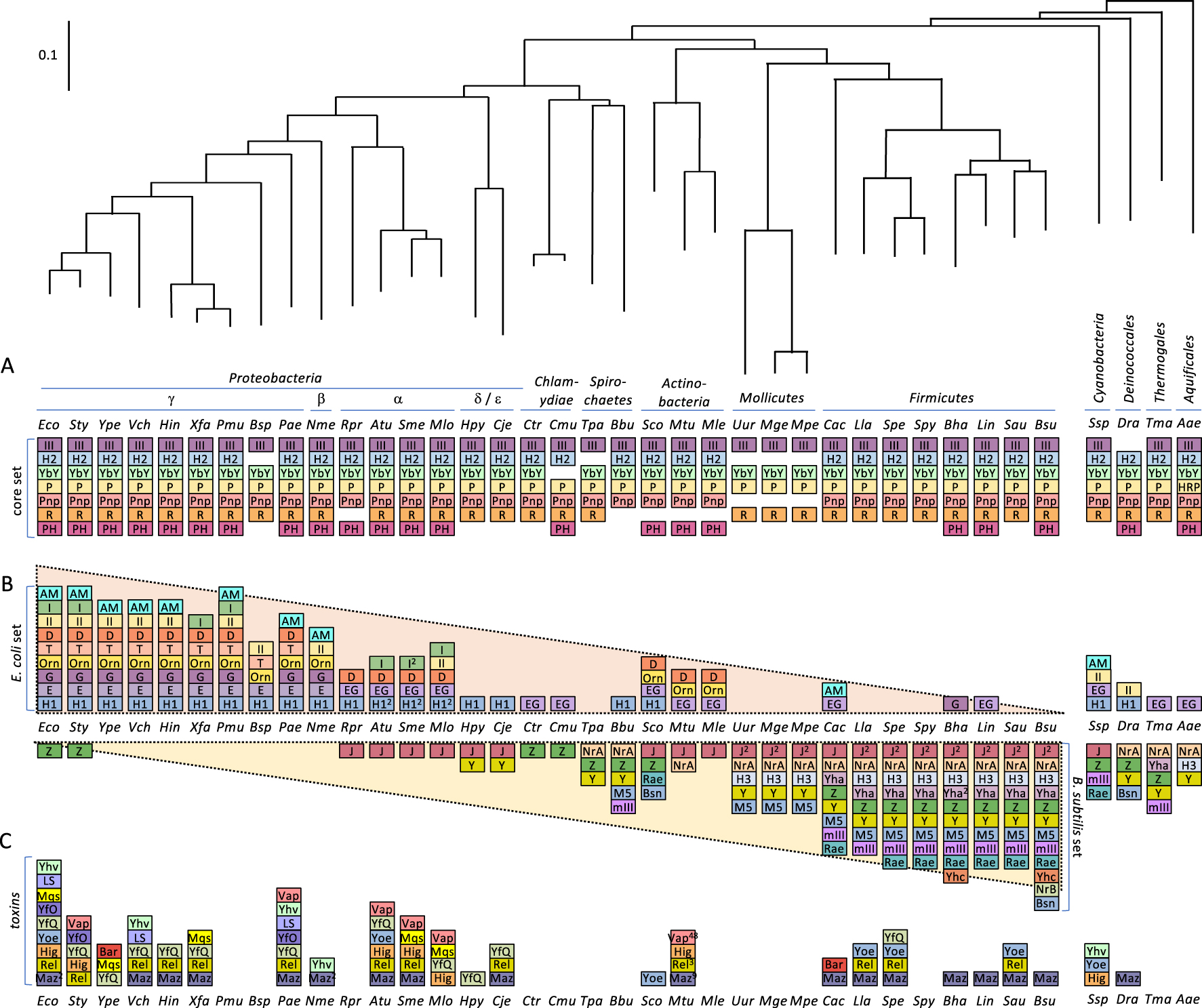
Remarkably, of the 38 RNases found in either E. coli and B. subtilis, only 9 are common to both (8 house-keeping RNases and 1 toxin). Seven of these house-keeping enzymes are highly conserved throughout bacteria and thus constitute a “core” set (Figure 1A). These include two tRNA maturation enzymes (RNase P and RNase PH), three rRNA maturation and degrading enzymes (RNase III, YbeY and RNase R), the mRNA degrading enzyme PNPase, and an enzyme that degrades the RNA primers used to initiate DNA replication, RNase H2. These enzymes are found in the vast majority bacteria except obligate intracellular parasites such as Mycoplasma, which have significantly reduced genomes, afforded by their relatively stable environments. In addition to the seven enzymes in the highly conserved “core set”, E. coli and B. subtilis have their own particular sets of house-keeping RNases that perform the tasks of RNA maturation and decay. These two organisms are about as far apart as it is possible to get on the bacterial phylogenetic tree (Figure 1). As one moves step by step along the tree from the γ-Proteobacteria through the α, β, and δ∕ε branches and beyond to the Chlamydiae, Spirochaetes, Actinobacteria, Mollicutes and eventually to the Firmicutes, the E. coli-specific set of RNases progressively fades and the B. subtilis set appears (Figure 1B). In the next few paragraphs, we will describe what these enzymes do in each organism and where they are primarily found in between.
1. Distribution of enzymes involved in mRNA degradation
E. coli and B. subtilis use overlapping strategies but very different enzymes to degrade their mRNAs. In E. coli, the key RNase that makes the first endoribonucleolytic cut is RNase E, a low specificity enzyme with a preference for AU-rich single stranded (ss) regions, with a strong preference for a U-residue at +2 relative to the cleavage site [12]. It is essential in E. coli and is associated with the inner membrane via an amphipathic helix downstream of the N-terminal catalytic half of the protein [13]. The C-terminal half is globally unstructured, but has structured “microdomains” that allow tethering of the RhlB helicase, the glycolytic enzyme enolase and the 3′–5′ exoribonuclease PNPase to create the canonical degradosome [14]. Although RNase E can cleave about half of its substrates internally without contacting the 5′ end [15], for the remaining substrates, cleavage efficiency is improved by about two orders of magnitude if the 5′ end of the mRNA contains a monophosphate group [16]. This can be the result of a prior endoribonucleolytic cleavage or removal of the 5′-triphosphate group typically present in primary transcripts by an RNA pyrophosphohydrolase such as RppH [17]. Most of the γ- and β-Proteobacteria have a paralog of RNase E called RNase G that has similar properties but that lacks the C-terminal degradosome domain. These two enzymes have clearly evolved from a duplication of a gene encoding an enzyme with similar levels of homology to both, known as RNase EG. RNase EG is widespread but is rarer in the Firmicutes and lacking in B. subtilis.
The initial endoribonucleolytic cleavage can also be performed by other enzymes such as the double strand (ds) specific RNase III, found in the core set. Endoribonucleolytic cleavage gives access to the 3′ end to a number of redundant 3′–5′ exoribonucleases, principally RNase II and PNPase, and occasionally RNase R and RNase PH. All but RNase II are found in the core set of RNases. RNase II, which likely evolved from a duplication of the RNase R gene, is well conserved in the γ- and β-Proteobacteria, but only occurs sporadically outside of these two classes. All of these 3′ exoribonucleases typically cannot degrade RNAs all the way to mononucleotides and fall off the substrates when they are in the range of 2–10 nts [18]. The essential enzyme oligoribonuclease, encoded by the orn gene, converts these end-products to mononucleotides [19]. Like RNase E, RNase G and RNase II, oligoribonuclease primarily occurs in γ- and β-Proteobacteria, but also pops up in the Actinobacteria, thought to have resulted from the fusion of a Proteobacterium and a Firmicute genome.
In B. subtilis, the key endoribonuclease involved in initiating mRNA turnover is RNase Y [20, 21]. Although it has no homology to RNase E, it shares many of its properties including membrane localization (via an N-terminal transmembrane helix) [22], its loose specificity for single-stranded AU-rich sequence (but with cleavage preferentially after a G-residue) [23] and an ability to associate at least transiently with helicase CshA and PNPase, creating a degradosome like network [24, 25], whose functionality remains to be proven. The activity of RNase Y has also been shown to be stimulated by a 5′-monophosphate for at least one substrate [20]. RNase Y begins to appear in the δ∕ε-Proteobacteria and is widely conserved all the way through to the Firmicutes, and in the Deinoccales, Thermatogales and Aquificales branches of the phylogenetic tree. Although a number of organisms have both RNase EG and RNase Y, they generally tend to be mutually exclusive, as one might expect from their analogous functions. Indeed E. coli RNase E can functionally replace RNase Y in B. subtilis [26], a remarkable observation given the evolutionary distance between these two organisms and the complete lack of homology between the two enzymes.
As in E. coli, RNase III (core set) can also initiate decay by cleaving RNA in ds regions. In B. subtilis, the fragments generated by endoribonucleolytic cleavage can be degraded in either the 5′–3′ direction by RNase J1 [27], which forms a complex with its paralog RNase J2 [28], or in the 3′–5′ direction by redundant exoribonucleases PNPase, RNase R, YhaM and RNase PH, with PNPase thought to be the primary source of this activity [29]. The RNase J1/J2 complex that is absent in E.coli, can initiate mRNAs degradation directly from the 5′ end following prior removal of the 5′ triphosphate group by BsRppH [17]. Exoribonucleotic degradation from the 5′ end is a major pathway in B.subtilis that is similar to that found in eukaryotes, where decapped RNA is degraded by the 5′ exoribonuclease Xrn1. RNase J is highly conserved from the α-Proteobacteria all the way through to the Mollicutes and Firmicutes, where its gene was duplicated to produce RNase J1 and J2.
2. Distribution of enzymes involved in tRNA maturation
While both E. coli and B. subtilis use the ubiquitous ribozyme RNase P (core set) to mature the 5′ ends of their tRNAs, they show major differences in the enzymes used for the 3′ processing step. In E. coli, all tRNA genes have the 3′ CCA motif, to which the cognate amino acid is attached, directly encoded by their genes, whereas in B. subtilis, this is only the case for about one third of the 86 tRNAs encoded by the genome and the CCA motif is attached later by an enzyme called nucleotidyl transferase or CCase. In E. coli, tRNA 3′ maturation is catalysed by up to 5 redundant 3′ exoribonucleases RNase T, RNase PH (core set), RNase D, RNase II and RNase Z (also known as RNase BN) [30]. While RNase T is confined to the γ-Proteobacteria, RNase D is conserved as far as the α-class and occurs again in the Actinobacteria. RNase Z is widely conserved from the Chlamydiae to the Firmicutes, and in the Cyanobacterial, Deincoccales and Thermatogales branches, but is essentially absent from Proteobacteria, with the very curious exception of E. coli and Salmonella.
In B. subtilis, two tRNA maturation pathways are used, depending nominally on whether or not the CCA motif is encoded [31]. For those lacking the encoded CCA motif, processing is performed endoribonucleolytically by RNase Z at the first U residue that occurs downstream (0–2 nts) of the so-called discriminator nucleotide (nt 73) to which the CCA motif is added [31]. RNase Z (like RNase J1 and J2) belongs to the β-lactamase family of enzymes, that possess both endo- and exoribonuclease activity. Following endoribonucleolytic cleavage 5′ to the U residue, it uses its 3′ exoribonuclease activity to trim off any extra nts downstream of the discriminator nt. For those tRNAs with an encoded CCA motif, 3′ maturation occurs much as in E. coli, using a different set of redundant 3′ exoribonucleases, the core set RNases RNase PH, PNPase, RNase R and the Firmicute-confined YhaM. At least one other enzyme from this pathway remains to be identified.
Ribosomal RNA maturation enzymes in E. coli and B. subtilis. The pathways are shown on fully assembled ribosomes. Endoribonucleases are depicted as scissors symbols. Mature rRNA sequences are shown in red; precursor sequences in gold. For full description see text.
3. Distribution of enzymes involved in rRNA maturation
Perhaps the most spectacular contrast between E. coli and B. subtilis is found in the rRNA maturation pathways (Figure 2). Ribosomal RNAs (16S, 23S and 5S rRNAs) are transcribed in most bacteria as a ∼5 kb precursor (30S) that undergoes co-transcriptional processing by RNase III at two long ds processing stalks that flank the mature 16S and 23S rRNA sequences. Subsequent to this step E. coli 16S rRNA is matured at its 5′ end by the endoribonucleases RNases E and G [32] and the final 3 nts removed by the 5′ exoribonuclease RNase AM [33], primarily found in the γ- and β-Proteobacteria. This step is done by RNase J1/J2 in B. subtilis [34]. 3′ processing of 16S rRNA can proceed either exoribonucleolytically through PNPase, RNase R, RNase II, RNase PH [35] or endoribonucleolytically through the ubiquitous enzyme YbeY [36], a member of the core set and one of only about 200 proteins conserved in essentially all bacteria [37]. B. subtilis uses only the YbeY homolog YqfG for this step [38]. In E. coli 23S processing is performed by the exoribonucleases RNase AM and RNase T at the 5′ and 3′ ends, respectively [33, 39], while in B. subtilis these reactions are performed by the ds specific enzyme Mini-RNase III that matures the 5′ and 3′ ends together [40, 41]. Mini-III is primarily a Firmicute enzyme, only occurring sporadically in other organisms, notably the Spirochaetes, the Cyanobacteria and the Thermatogales. Lastly 5S rRNA is matured by RNase E followed by RNase AM (5′) and RNase T (3′) in E. coli [33, 42], whereas B. subtilis 5S rRNA is matured by the dedicated ds endoribonuclease RNase M5 [40, 43], mostly confined to the Mollicutes and Firmicutes. Remarkably, only the YbeY-catalysed 16S rRNA final maturation step is conserved between E. coli and B. subtilis and most of the key enzymes used for rRNA processing in E. coli (RNases E, G, AM, T) are not found in B. subtilis and vice versa (RNases J1, J2, Mini-III, M5). Thus, despite the high level of conservation of the rRNAs and r-proteins that make up the ribosome, these two bacteria have evolved with remarkably different maturation machineries. These processing reactions are different again in the archaea and in eukaryotes suggesting that rRNA processing pathways have been invented at least four times in nature.
It is clear that many more bacterial RNases remain to be discovered, in particular among the toxin class of RNases that has been the major source of new enzymes in recent times. It is likely that organisms lying in the furthest branches of the phylogenetic tree from the two model organisms discussed here (e.g. the Cyanobacteria, Deincoccales, Thermatogales and Aquificales) will be the richest sources of new housekeeping enzymes to degrade and mature bacterial RNAs.
Conflicts of interest
Authors have no conflict of interest to declare.
Acknowledgments
This work was supported by Centre National de la Recherche Scientifique (CNRS) and the Université de Paris to UMR8261, the Agence Nationale de la Recherche (CoNoCo, BSpoRNase) and the Labex Dynamo program.
French version
Lorsque la séquence du génome de Bacillus subtilis a été achevée en 1997, son exhaustivité a été remise en question par des collègues scientifiques qui avaient du mal à accepter l’absence du gène codant pour la ribonucléase essentielle et la plus importante chez Escherichia coli en termes de nombre de substrats (RNase E). En effet, la séquence complète du génome a permis de réaliser que non seulement le gène codant pour la RNase E (rne) était absent de B. subtilis, mais que sur les ∼15 ribonucléases connues chez E. coli à l’époque, seules quelques-unes avaient des homologues chez B. subtilis (RNase P, RNase III, RNase PH, PNPase et RNase R). Cela a ouvert un tout nouveau champ de recherche pour notre laboratoire, et pour d’autres, en essayant de trouver les composants manquants de la machinerie de maturation et de dégradation de l’ARN de B. subtilis. Aujourd’hui, B. subtilis a été clairement établi comme un modèle alternatif au paradigme d’E. coli pour le métabolisme des ARN. Dans cette revue, nous décrirons ces deux extrémités du spectre phylogénétique et montrerons comment deux gradients opposés, représentant l’apparition ou la disparition de ribonucléases spécifiques, peuvent expliquer la grande majorité de l’espace évolutif entre ces deux organismes qui se sont séparés il y a 1 à 3 milliards d’années.
La quantité de tout ARN dans la cellule et donc potentiellement la quantité de protéine qu’il code est un équilibre entre son taux de synthèse et de dégradation. Si les mécanismes de transcription et de traduction sont remarquablement bien conservés chez les bactéries, ce n’est pas le cas du mécanisme de dégradation de l’ARN. Chez les bactéries, l’ARN est dégradé par une combinaison d’endo- et d’exo-ribonucléases qui coupent l’ARN de façon interne ou retirent un nucléotide (nt) à la fois de l’extrémité 5′ ou 3′. L’activité de la RNase peut être modulée à la fois au niveau du substrat (par exemple, par des changements de conformation de sa structure secondaire ou tertiaire), ou par la présence de certains groupes chimiques à l’extrémité 5′ (par exemple, les groupes triphosphate 5′, les coiffes NAD, les coiffes NpnN, les coiffes CoA) ou à l’intérieur (par exemple, les modifications m6-A, pseudouridine (𝜓)) [1, 2, 3, 4, 5]. Elle peut également être modulée par des cofacteurs protéiques ou par des modifications post-traductionnelles (par exemple, acétylation, phosphorylation) [6, 7]. En plus de dégrader l’ARN, les RNases peuvent aussi paradoxalement stabiliser l’ARN, en éliminant les nucléotides qui pourraient potentiellement recruter des enzymes de dégradation. C’est typiquement le cas pour les ARN dits stables, par exemple l’ARN ribosomique (ARNr) et l’ARN de transfert (ARNt), qui sont généralement synthétisés sous forme de précurseurs qui sont coupés à leurs extrémités 5′ et 3′ pour donner les molécules finales stables et fonctionnelles [8]. Une même RNase peut avoir à la fois des fonctions de stabilisation (maturation/« processing ») et de dégradation selon le substrat de l’ARN et selon que son produit devient plus ou moins sensible aux autres RNases.
On connaît actuellement environ 40 RNases bactériennes, dont la grande majorité a été découverte chez E. coli ou B. subtilis. E. coli compte actuellement 27 RNases connues, tandis que B. subtilis en compte 21. La plupart d’entre elles sont impliquées dans des opérations dites de « ménage » telles que la dégradation de l’ARNm et la maturation de l’ARN stable, et leurs fonctions sont résumées dans le Tableau 1. Plusieurs appartiennent à la famille des RNases dites toxines. En effet, la principale différence dans le nombre total de RNases entre E. coli et B. subtilis est que E. coli possède au moins dix RNases toxines connues [9], alors que B. subtilis n’en possède qu’une seule à ce jour [10]. L’expression des paires toxine-antitoxine (TA) est généralement induite dans différentes conditions de stress et la libération de la toxine d’un complexe inhibiteur avec son antitoxine repose sur la dégradation rapide de l’antitoxine. Une fois que la toxine est libérée, elle agit comme une endoribonucléase, clivant l’ARN d’une manière spécifique à la séquence, typiquement avec des séquences de reconnaissance courtes de 4–6 nts, qui apparaissent très fréquemment. On pense que les modules TA sont transférés horizontalement et sont souvent associés à des éléments mobiles, dont certains sont devenus cryptiques avec le temps.
Ribonucléases retrouvées ches les bactéries
Enzyme type | Principle function(s) | |
|---|---|---|
| Core set | ||
| RNase III | Endoribonuclease | Cleavage of double stranded RNA: rRNA precursors, mRNA, antisense RNA |
| RNase H2 | Endoribonuclease | Cleavage of DNA/RNA hybrids |
| YbeY | Endoribonuclease | Processing of 16S rRNA 3′ end |
| RNase P | Endoribonuclease | Processing of tRNA 5′ ends |
| PNPase | 3′-exoribonuclease | Phosphorolytic degradation of mRNA, rRNA. Protection of sRNA |
| RNase R | 3′-exoribonuclease | Hydrolytic degradation of rRNA, mRNA |
| RNase PH | 3′-exoribonuclease | Phosphorolytic processing of tRNA 3′ ends |
| « E. coli » set | ||
| RNase E | Membrane associated endoribonuclease | Cleavage of mRNA, sRNA, processing of tRNA, 16S and 5S rRNA |
| RNase G | Endoribonuclease | Cleavage of mRNA, processing 16S rRNA |
| RNase H1 | Endoribonuclease | Cleavage of DNA/RNA hybrids |
| RNase I | Endoribonuclease | Periplasmic RNase, degradation of rRNA following membrane stress |
| Oligoribonuclease | 3′-exoribonuclease | Degradation of short oligonucleotides, end products of other 3′–5′ exoribonucleotides |
| RNase T | 3′-exoribonuclease | Hydrolytic processing of tRNA 3′ ends |
| RNase D | 3′-exoribonuclease | Hydrolytic processing of tRNA 3′ ends |
| RNase II | 3′-exoribonuclease | Hydrolytic processing of tRNA 3′ ends, shortening of mRNA 3′ ends |
| RNase AM | 5′-exoribonuclease | 5′ processing of 16S, 23S and 5S rRNA |
| « B. subtilis » set | ||
| RNase Y | Membrane associated endoribonuclease | Cleavage of mRNA, processing of RNase P RNA, sRNA |
| RNase Z | Endoribonuclease/3′-exoribonuclease | 3′ processing of tRNA |
| RNase H3 | Endoribonuclease | Cleavage of DNA/RNA hybrids |
| RNase M5 | Endoribonuclease | Processing of 5S rRNA |
| Mini-RNase III | Endoribonuclease | Processing of 23S rRNA, type II self-cleaving ribosymes |
| Rae1 | Endoribonuclease | Cleavage of mRNA through association with ribosome |
| YhcR | Membrane associated endoribonuclease | unknown |
| Bsn | Endoribonuclease | Degradation of extracellular RNA |
| RNase J | 5′–3′ exoribonuclease/ endoribonuclease | Degradation of mRNA, sRNA, processing of 16S rRNA 5′ end |
| NanoRNase A | 3′–5′ exoribonuclease | Degradation of short oligonucleotides, end products of other 3′–5′ exoribonucleotides |
| NanoRNase B | 3′–5′ exoribonuclease | Degradation of short oligonucleotides, end products of other 3′–5′ exoribonucleotides |
| YhaM | 3′–5′ exoribonuclease | 3′ processing of tRNA, shortening of mRNA 3′ ends |
Phylogeny of ribonucleases in bacteria. This is an update and new representation of Ref. [11] showing the first 38 model organisms sequenced, a measure of their scientific importance. (A) The « core set » of RNases conserved in most bacteria. (B) Distribution of the « E. coli set » and « B. subtilis set » of RNases across the phylogenetic tree. (C) Distribution of horizontally transferred toxin RNases across the phylogenetic tree. RNase abbreviations are as follows: III = RNase III; P = RNase P; H2 = RNase HII; H1 = RNase HI; H3 = RNase HIII; E = RNase E; G = RNase G; EG = RNase E/G; YbY = YbeY; I = RNase I; Z = RNase Z; M5 = RNase M5; Y = RNase Y; mIII = Mini-III; Rae = Rae1; Maz = MazF; Rel = RelE; Yoe = YoeB; Hig = HigB; YfQ = YafQ; Yhv = YhaV; LS = RNase LS, Mqs = MqsR; YfO = YafO; Bsn = RNase Bsn; Bar = Barnase; AM = RNase AM; R = RNase R; Pnp = polynucleotide phosphorylase; PH = RNase PH; D = RNase D; Orn = oligoribonuclease; II = RNase II; T = RNase T; J = RNase J; BN = RNase BN; Yha = RNase YhaM. Numbers in superscript refer to the number of orthologs present when identified. Organism abbreviations are as follows: Eco = Escherichia coli; Sty = Salmonella typhimurium; Ype = Yersinia pestis; Vch = Vibrio cholera; Hin = Haemophilus influenza; Xfa = Xylella fastidiosis; Pmu = Pasteurella multocida; Bsp = Buchnera sp.; Pae = Pseudomonas aeruginosa; Nme = Neisseria meningitidis; Rpr = Rickettsia prowazekii; Atu = Agrobacterium tumefaciens; Sme = Sinorhizobium meliloti; Mlo =Mesorhizobium loti; Hpy Helicobacter pylori; Cje =Campylobacter jejuni; Ctr =Chlamydia trachomatis; Cmu = Chlamydia muridarum; Tpa = Treponema pallidum; Bbu = Borrelia burgdorferi; Sco = Streptomyces coelicolor; Mtu = Mycobacterium tuberculosis; Mle = Mycobacterium leprae; Uur = Ureaplasma urealyticum; Mge = Mycoplasma genitalium; Mpe = M. pneumoniae; Cac = Clostridium acetobutylicum; Lla = Lactococcus lactis; Spe = Streptococcus pneumoniae; Spy = Streptococcus pyogenes; Bha = Bacillus halodurans; Lin = Listeria innocua; Sau = Staphylococcus aureus; Bsu = Bacillus subtilis; Ssp = Synechocystis sp.; Dra = Deinococcus radiodurans; Tma = Thermotoga maritima; Aao = Aquifex aeolicus.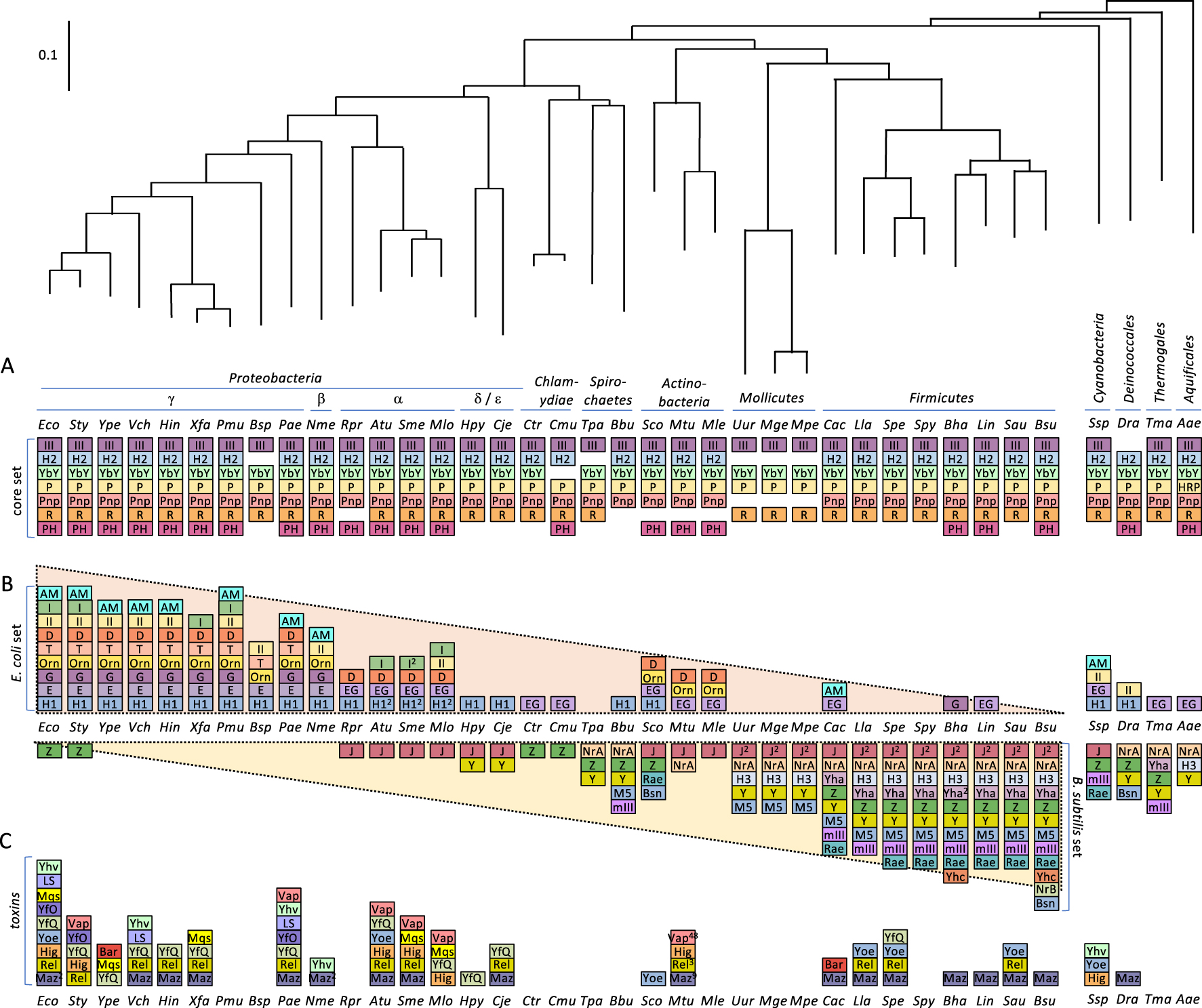
Il est remarquable de constater que sur les 38 RNases présentes chez E. coli et B. subtilis, seules 9 sont communes aux deux (8 RNases de ménage et 1 toxine). Sept de ces enzymes de ménage sont hautement conservées dans l’ensemble des bactéries et constituent donc un ensemble « coeur » (Figure 1A). Celles-ci incluent deux enzymes de maturation d’ARNt (RNase P et RNase PH), trois enzymes de maturation et de dégradation d’ARNr (RNase III, YbeY et RNase R), l’enzyme de dégradation d’ARNm PNPase, et une enzyme qui dégrade les amorces d’ARN utilisées pour initier la réplication d’ADN, RNase H2. Ces enzymes sont présentes dans la grande majorité des bactéries, à l’exception des parasites intracellulaires obligatoires tels que les mycoplasmes, qui ont des génomes considérablement réduits, permis par leur environnement relativement stable. Outre les sept enzymes du « noyau » hautement conservé, E. coli et B. subtilis possèdent leurs propres ensembles de RNases de ménage qui assurent la maturation et la dégradation de l’ARN. Ces deux organismes sont à peu près aussi éloignés l’un de l’autre qu’il est possible de l’être sur l’arbre phylogénétique bactérien (Figure 1). En se déplaçant pas à pas le long de l’arbre, des γ-Proteobacteria aux branches α, β et δ∕ε, puis aux Chlamydiae, Spirochaetes, Actinobacteria, Mollicutes et finalement aux Firmicutes, l’ensemble des RNases spécifiques à E. coli s’estompe progressivement et l’ensemble de B. subtilis apparaît (Figure 1B). Dans les prochains paragraphes, nous décrirons ce que font ces enzymes dans chaque organisme et où on les trouve principalement entre les deux.
1. Distribution des enzymes impliquées dans la dégradation des ARNm
E. coli et B. subtilis utilisent des stratégies qui se chevauchent mais des enzymes très différentes pour dégrader leurs ARNm. Chez E. coli, la RNase clé qui effectue la première coupure endoribonucléolytique est la RNase E, une enzyme de faible spécificité avec une préférence pour les régions simple-brins (ss) riches en AU, avec une forte préférence pour un résidu U à +2 par rapport au site de coupure [12]. Elle est essentielle chez E. coli et est associée à la membrane interne par une hélice amphipathique située en aval de la moitié catalytique N-terminale de la protéine [13]. La moitié C-terminale est globalement non structurée, mais possède des « microdomaines » structurés qui permettent d’attacher l’hélicase RhlB, l’enzyme glycolytique énolase et l’exoribonucléase 3′–5′ PNPase pour créer le dégradosome canonique [14]. Bien que la RNase E puisse cliver environ la moitié de ses substrats en interne sans entrer en contact avec l’extrémité 5′ [15], pour les substrats restants, l’efficacité du clivage est améliorée d’environ deux ordres de grandeur si l’extrémité 5′ de l’ARNm contient un groupe monophosphate [16]. Cela peut être le résultat d’un clivage endoribonucléolytique préalable ou de l’élimination du groupe 5′-triphosphate généralement présent dans les transcrits primaires par une ARN pyrophosphohydrolase telle que RppH [17]. La plupart des γ- et β-Protéobactéries possèdent un paralogue de la RNase E appelé RNase G qui possède des propriétés similaires mais qui est dépourvu du domaine dégradosome C-terminal. Ces deux enzymes ont clairement évolué à partir d’une duplication d’un gène codant pour une enzyme présentant des niveaux d’homologie similaires aux deux, connue sous le nom de RNase EG. La RNase EG est très répandue mais est plus rare chez les Firmicutes et absente chez B. subtilis.
Le clivage endoribonucléolytique initial peut également être effectué par d’autres enzymes, comme la RNase III spécifique au double brin (ds), que l’on trouve dans l’ensemble coeur. Le clivage endoribonucléolytique donne accès à l’extrémité 3′ à un certain nombre d’exoribonucléases 3′–5′ redondantes, principalement la RNase II et la PNPase, et occasionnellement la RNase R et la RNase PH. Toutes ces exoribonucléases, à l’exception de la RNase II, font partie de l’ensemble cœur des RNases. La RNase II, qui a probablement évolué à partir d’une duplication du gène de la RNase R, est bien conservée dans les γ- et β-Proteobacteria, mais n’est présente que sporadiquement en dehors de ces deux classes. Toutes ces exoribonucléases 3′ ne peuvent généralement pas dégrader les ARN jusqu’aux mononucléotides et relarguent leurs substrats lorsqu’ils se situent entre 2 et 10 nts [18]. L’enzyme essentielle l’oligoribonucléase, codée par le gène orn, convertit ces produits finaux en mononucléotides [19]. Comme la RNase E, la RNase G et la RNase II, l’oligoribonucléase est principalement présente chez les γ- et β-Protéobactéries, mais elle apparaît également chez les Actinobactéries, que l’on pense être le résultat de la fusion d’un génome de Protéobactérie et de Firmicute.
Chez B. subtilis, l’endoribonucléase clé impliquée dans l’initiation du renouvellement de l’ARNm est la RNase Y [20, 21]. Bien qu’elle n’ait pas d’homologie avec la RNase E, elle partage beaucoup de ses propriétés, y compris la localisation membranaire (via une hélice transmembranaire N-terminale) [22], sa spécificité pour la séquence riche en AU simple brin (mais avec un clivage préférentiel après un résidu G) [23] et une capacité à s’associer au moins transitoirement avec l’hélicase CshA et la PNPase, créant un réseau semblable à un dégradosome [24, 25], dont la fonctionnalité reste à prouver. Il a également été démontré que l’activité de la RNase Y est stimulée par un 5′-monophosphate pour au moins un substrat [20]. La RNase Y commence à apparaître chez les δ∕ε-Proteobacteria et est largement conservée jusqu’aux Firmicutes, et dans les branches Deinoccales, Thermatogales et Aquificales de l’arbre phylogénétique. Bien qu’un certain nombre d’organismes possèdent à la fois la RNase EG et la RNase Y, elles ont généralement tendance à s’exclure mutuellement, comme on pourrait s’y attendre en raison de leurs fonctions analogues. En effet, la RNase E d’E. coli peut remplacer fonctionnellement la RNase Y de B. subtilis [26], une observation remarquable étant donné la distance évolutive entre ces deux organismes et l’absence totale d’homologie entre les deux enzymes.
Comme chez E. coli, la RNase III (ensemble coeur) peut également initier la dégradation en clivant l’ARN dans les régions ds. Chez B. subtilis, les fragments générés par le clivage endoribonucléolytique peuvent être dégradés soit dans la direction 5′–3′ par la RNase J1 [27], qui forme un complexe avec sa paralogue la RNase J2 [28], soit dans la direction 3′–5′ par les exoribonucléases redondantes PNPase, RNase R, YhaM et RNase PH, la PNPase étant considérée comme la source principale de cette activité [29]. Le complexe RNase J1/J2, absent chez E.coli, peut initier la dégradation des ARNm directement à partir de l’extrémité 5′ après élimination préalable du groupe triphosphate 5′ par BsRppH [17]. La dégradation exoribonucléotique depuis l’extrémité 5′ est une voie majeure chez B.subtilis qui est similaire à celle trouvée chez les eucaryotes, où l’ARN décoiffé est dégradé par l’exoribonucléase 5′ Xrn1. La RNase J est hautement conservée depuis les α-protéobactéries jusqu’aux Mollicutes et Firmicutes, où son gène a été dupliqué pour produire la RNase J1 et J2.
2. Distribution des enzymes impliquées dans la maturation des ARNt
Alors que E. coli et B. subtilis utilisent tous deux la ribozyme omniprésente RNase P (ensemble coeur) pour maturer les extrémités 5′ de leurs ARNt, ils montrent des différences majeures dans les enzymes utilisées pour l’étape de maturation 3′. Chez E. coli, tous les gènes d’ARNt ont le motif 3′ CCA, auquel l’acide aminé correspondant est attaché, directement codé par leurs gènes, alors que chez B. subtilis, ce n’est le cas que pour environ un tiers des 86 ARNt codés par le génome et le motif CCA est attaché plus tard par une enzyme appelée nucléotidyl transférase ou CCase. Chez E. coli, la maturation en 3′ des ARNt est catalysée par jusqu’à 5 exoribonucléases 3′ redondantes : la RNase T, la RNase PH (ensemble de base), la RNase D, la RNase II et la RNase Z (également appelée RNase BN) [30]. Alors que la RNase T est confinée aux γ-Proteobacteria, la RNase D est conservée jusqu’à la classe α et se retrouve chez les Actinobacteria. La RNase Z est largement conservée des Chlamydiae aux Firmicutes, et dans les branches des Cyanobactéries, des Deincoccales et des Thermatogales, mais est essentiellement absente des Protéobactéries, à l’exception très curieuse d’E. coli et de Salmonella.
Chez B. subtilis, deux voies de maturation des ARNt sont utilisées, selon que le motif CCA est codé ou non, à priori [31]. Pour ceux qui ne possèdent pas le motif CCA codé, la maturation est effectuée de manière endoribonucléolytique par la RNase Z au niveau du premier résidu U qui se trouve en aval (0–2 nts) du nucléotide dit discriminant (nt 73) auquel le motif CCA est ajouté [31]. La RNase Z (comme la RNase J1 et J2) appartient à la famille des β-lactamases, qui possèdent une activité endo- et exoribonucléase. Après le clivage endoribonucléolytique en 5′ du résidu U, elle utilise son activité exoribonucléase en 3′ pour enlever tout nt supplémentaire en aval du nt discriminant. Pour les ARNt ayant un motif CCA codé, la maturation en 3′ se produit à peu près comme chez E. coli, en utilisant un ensemble différent d’exoribonucléases 3′ redondantes, les RNases de l’ensemble cœur, la RNase PH, la PNPase et la RNase R, ainsi que la RNase YhaM confinée aux Firmicutes. Au moins une autre enzyme de cette voie reste à identifier.
Enzymes de maturation de l’ARN ribosomique chez E. coli et B. subtilis. Les voies sont montrées sur les ribosomes complètement assemblés. Les endoribonucléases sont représentées par des symboles de ciseaux. Les séquences d’ARNr matures sont représentées en rouge ; les séquences précurseurs en or. Pour une description complète, voir le texte.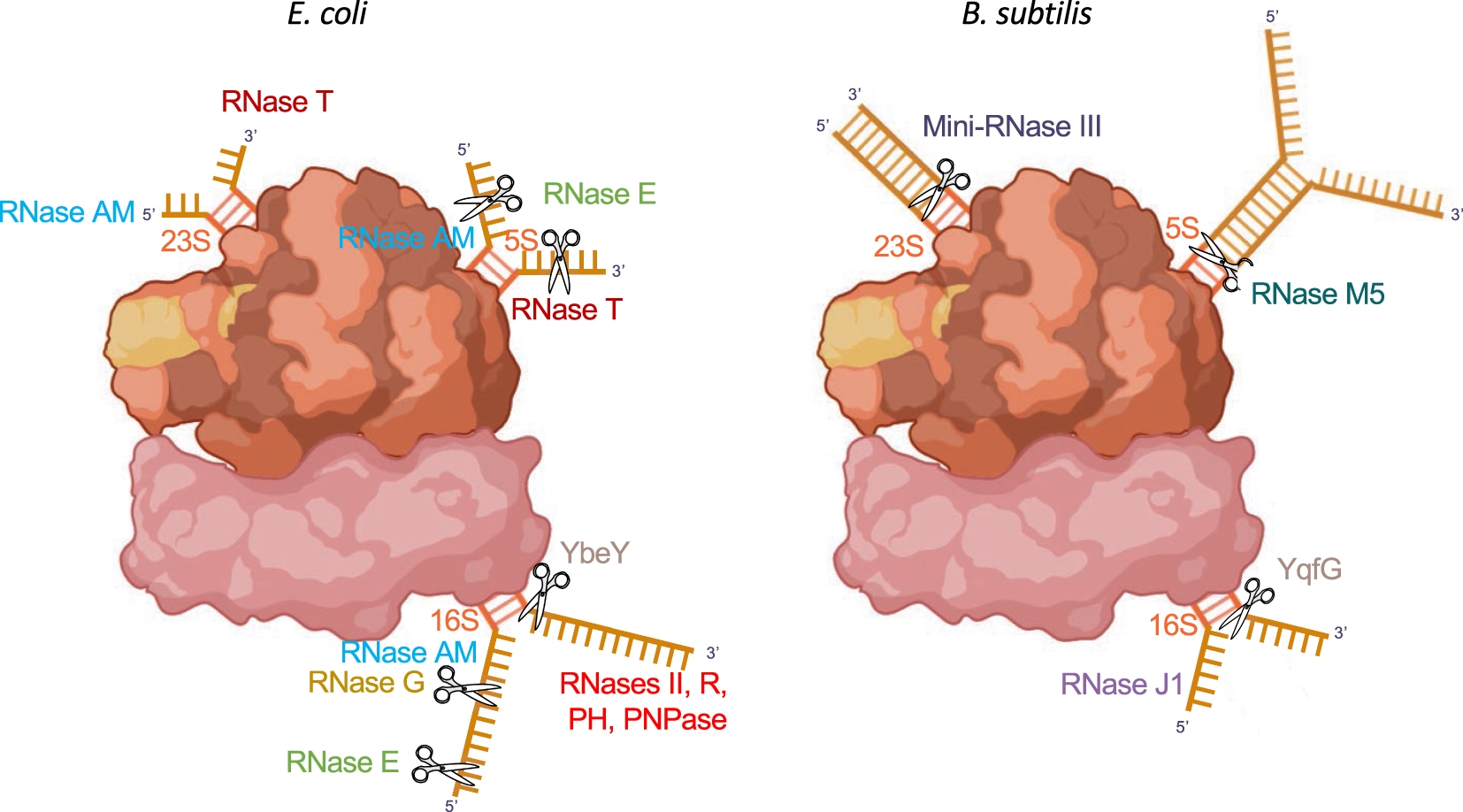
3. Distribution des enzymes impliquées dans la maturation des ARNr
Le contraste le plus spectaculaire entre E. coli et B. subtilis se trouve peut-être dans les voies de maturation des ARNr (Figure 2). Les ARN ribosomiques (ARNr 16S, 23S et 5S) sont transcrits dans la plupart des bactéries sous la forme d’un précurseur de ∼5 kb (30S) qui subit une maturation co-transcriptionnelle par la RNase III au niveau de deux longues tiges de transformation ds qui flanquent les séquences d’ARNr 16S et 23S matures. Après cette étape, l’ARNr 16S d’E. coli est maturé à son extrémité 5′ par les endoribonucléases RNases E et G [32] et les 3 derniers nts sont éliminés par la 5′ exoribonucléase, la RNase AM [33], que l’on trouve principalement chez les γ- et β-protéobactéries. Cette étape est réalisée par la RNase J1/J2 chez B. subtilis [34]. La maturation en 3′ de l’ARNr 16S peut se faire soit de manière exoribonucléolytique par la PNPase, la RNase R, la RNase II, la RNase PH [35], soit de manière endoribonucléolytique par l’enzyme ubiquitaire YbeY [36], un membre de l’ensemble coeur et l’une des 200 protéines conservées dans pratiquement toutes les bactéries [37]. B. subtilis utilise uniquement l’homologue de YbeY, YqfG, pour cette étape [38]. Chez E. coli, la maturation du 23S est effectuée par les exoribonucléases RNase AM et RNase T aux extrémités 5′ et 3′, respectivement [33, 39], tandis que chez B. subtilis, ces réactions sont effectuées par l’enzyme spécifique du de l’ARN ds, la Mini-RNase III, qui clive les extrémités 5′ et 3′ ensemble [40, 41]. La Mini-RNase III est principalement une enzyme des Firmicutes, n’apparaissant que sporadiquement dans d’autres organismes, notamment les Spirochaetes, les Cyanobactéries et les Thermatogales. Enfin l’ARNr 5S est maturé par la RNase E, suivie par la RNase AM (5′) et la RNase T (3′) dans E. coli [33, 42], alors que l’ARNr 5S de B. subtilis est maturé par l’endoribonucléase ds dédiée, la RNase M5 [40, 43], principalement confiné aux Mollicutes et Firmicutes. Il est remarquable que seule l’étape de maturation finale de l’ARNr 16S catalysée par YbeY soit conservée entre E. coli et B. subtilis et que la plupart des enzymes clés utilisées pour la maturation de l’ARNr chez E. coli (RNases E, G, AM, T) ne se retrouvent pas chez B. subtilis et vice versa (RNases J1, J2, Mini-III, M5). Ainsi, malgré le haut niveau de conservation des ARNr et des r-protéines qui composent le ribosome, ces deux bactéries ont évolué avec des machineries de maturation remarquablement différentes. Ces réactions de maturation sont encore différentes chez les archées et chez les eucaryotes, ce qui suggère que les voies de maturation des ARNr ont été inventées au moins quatre fois dans la nature.
Il est clair que de nombreuses autres RNases bactériennes restent à découvrir, en particulier parmi la classe des toxines RNases qui a été la principale source de nouvelles enzymes ces derniers temps. Il est probable que les organismes situés dans les branches les plus éloignées de l’arbre phylogénétique des deux organismes modèles discutés ici (par exemple les Cyanobactéries, les Deincoccales, les Thermatogales et les Aquificales) seront les sources les plus riches de nouvelles enzymes de ménage pour dégrader et maturer les ARN bactériens.
Conflit d’intérêt
Les auteurs n’ont aucun conflit d’intérêt à déclarer.
Remerciements
Ce travail a été soutenu par le Centre National de la Recherche Scientifique (CNRS) et l’Université de Paris à l’UMR8261, l’Agence Nationale de la Recherche (CoNoCo, BSpoRNase) et le programme Dynamo du Labex.