

1 Introduction
The principle of competitive exclusion [13,17,20,35,47] asserts that two or more competitors cannot coexist indefinitely on a single prey, which was supported by experiments on Paramecium cultures by Gause [15] (see also [28]). It was thought to hold in laboratory settings until Ayala [2] demonstrated experimentally that two species of Drosophila could coexist upon a single prey.
In order to explain Ayala's experiments, various competition models have been proposed. For example, Armstrong and McGehee [1] considered the model:
| (1.1) |
| (1.2) |
Consider an ordinary differential equation model for n interacting biological species:
| (1.3) |
To have the strong version of coexistence, researchers have taken various other factors into account when modeling competition, such as interspecific interference [45,46], spatial heterogeneity [5], stoichiometric principles [34], etc. Another important factor is intraspecific interference within a population of competitors, which includes aggressive displays, posturing, fighting, infanticide, and cannibalism [30]. When the principal effect of the intraspecific interference is a reduced rate of feeding or resource intake, the effects can be modeled via an altered functional response for the consumer [8,10,40]. Recently, Cantrell et al. [6] extended system (1.2) to incorporate conspecific feeding interference for one of the competitors. The functional responses for the competitor w and the prey u are now taken to have the Beddington–DeAngelis form:
On the other hand, when lethal fighting or cannibalism occurs, it is more appropriate to include a nonlinear (density-dependent) mortality term for the consumer into the competition model [30]. In plant population, density-dependent mortality is one of the three principal effects resulting from intraspecific competition [38,49]. To incorporate such intraspecific competition (as well as the interspecific competition) into the model, Ruan and He [39] studied the global stability of a chemostat-type competition model, Kuang et al. [30] investigated the global stability of a Lotka–Volterra competition model. See also [16,32,33] for persistence of n species on a single resource.
Notice that in Refs. [16,30,32,33,39] all competitors are assumed to have density-dependent mortality rates. A very natural and very significant question arises: If one of the competitors does not have a density-dependent mortality rate, does the model still exhibit coexistence and have a stable componentwise positive equilibrium? The numerical simulations of Kuang et al. [30] indicate that, even for a Lotka–Volterra model, a competitor without a density-dependent mortality rate could eliminate another competitor with a density-dependent mortality rate if the first competitor has a lower break-even prey biomass. The problem can be very subtle.
Hixon and Jones [21] found that density-dependent mortality in demersal marine fishes is often caused by the interplay of predation and competition (see also [22]). To study how the nonlinear mortality rate determines the dynamics of such competition models qualitatively, we consider a two-competitor/one-prey model of the form:
| (1.4) |
| (1.5) |
We shall show that the two competitors can coexist upon the single prey. It demonstrates that density-dependent mortality in one of the competitors can prevent competitive exclusion.
As an example, we apply the obtained results to a two-competitor/one-prey model with a Holling II functional response:
| (1.6) |
2 Mathematical analysis
2.1 General functional responses
First of all, we can see that the solutions to the initial value problem (1.4)–(1.5) are nonnegative.
Define
Next, we consider the existence of a positive equilibrium. System (1.4) has a componentwise positive equilibrium
| (2.1) |
| (2.2) |
| (2.3) |
It is important to note that the key assumption for the existence of the positive equilibrium
Now we study the local stability of the positive equilibrium
| (2.4) |
| (2.5) |
| (2.6) |
Since
- (i)
- (ii)
| (2.7) |
| (2.8) |
Theorem 2.1
Assume that the positive equilibrium
| (2.9) |
| (2.10) |
Remark 2.2
The conditions (2.9) and (2.10) (corresponding (2.17) and (2.18) in Proposition 2.4) are technical assumptions on parameters. For the specific Holling II functional response, these conditions will be expressed explicitly, see Remark 2.7. Also, since
Remark 2.3
Assume
Finally, we discuss the global stability of the positive equilibrium
| (2.11) |
In next subsection, for the model with Holling II functional responses, we will choose proper
2.2 Holling II functional responses
In this subsection we apply the above results to system (1.4) with Holling II functional responses, namely system (1.6). System (1.6) has a unique componentwise positive equilibrium
| (2.12) |
| (2.13) |
| (2.14) |
Define
| (2.15) |
| (2.16) |
By Theorem 2.1, we have the following local stability result.
Proposition 2.4
Assume that the positive equilibrium
| (2.17) |
| (2.18) |
Remark 2.5
Note that
Finally, we give a sufficient condition for the global stability of the positive equilibrium
Proposition 2.6
Assume that the positive equilibrium
| (2.19) |
Proof
Let
Remark 2.7
Using (2.12), (2.15) and (2.19), one of the local stability conditions (2.17) becomes:
| (2.20) |
| (2.21) |
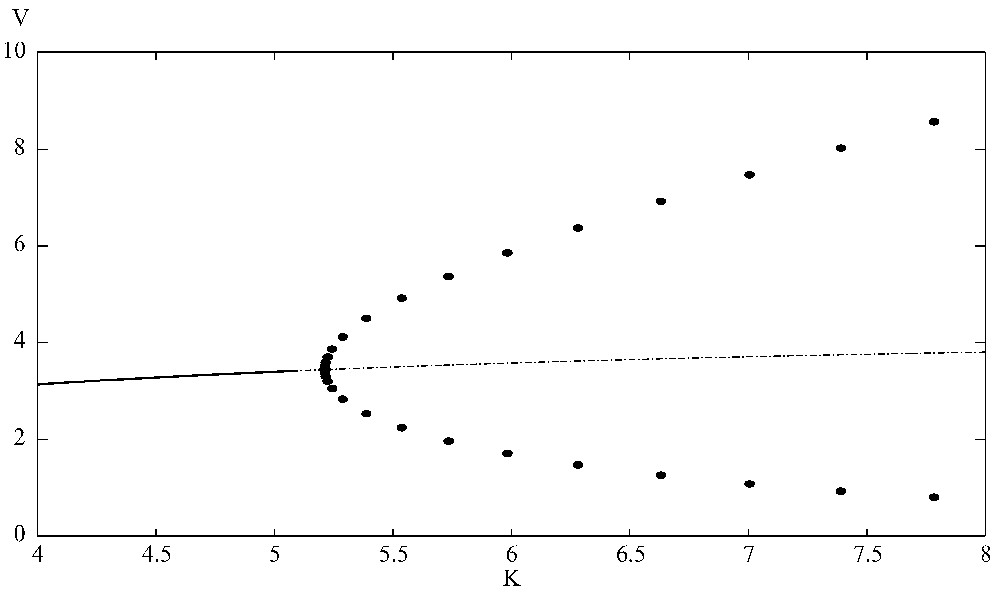
The bifurcation diagram shows that the positive equilibrium is stable when K⩽5.21. At K=5.21, it loses its stability and a supercritical Hopf bifurcation occurs. Here r=1.5, a=0.45, b=0.35, d=0.45, e=0.55, A=0.55, B=0.35, D=0.45, E=0.65, G=0.1. XPPAUT was used for the simulations.
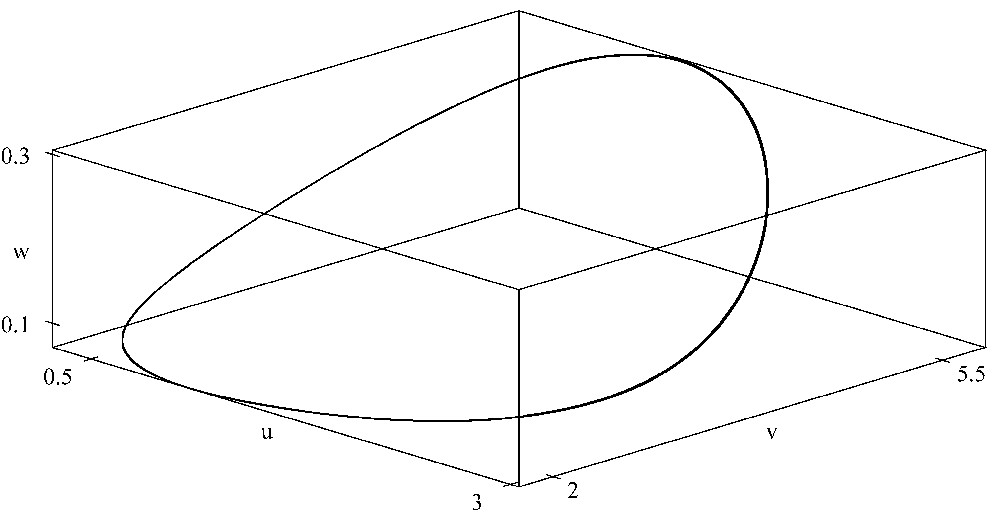
When K=6, there is a periodic orbit bifurcated from the interior equilibrium in the three-dimensional space which is asymptotically stable. Here r=1.5, a=0.45, b=0.35, d=0.45, e=0.55, A=0.55, B=0.35, D=0.45, E=0.65, G=0.1. XPPAUT was used for the simulations.
The plot of
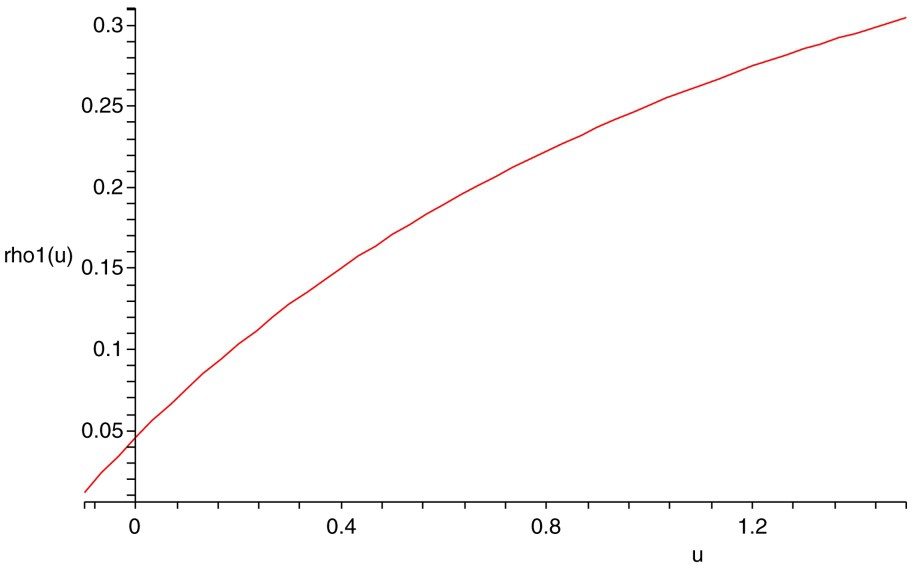
The plot of the function
3 Simulations and discussion
It is well known that density-dependent mortality terms (closure terms) can greatly affect the outcome of plankton models [42]: not only limit cycles [11] but also chaos [7] can occur in such models.
In the case of two competitors competing for a common prey, our results indicate that density-dependent mortality of one competitor not only ensures the long-term survival of itself, but also guarantees the existence of the other competitor, which would otherwise be out-competed.
To illustrate the results numerically, consider system (1.6) with Holling II functional responses. Choose
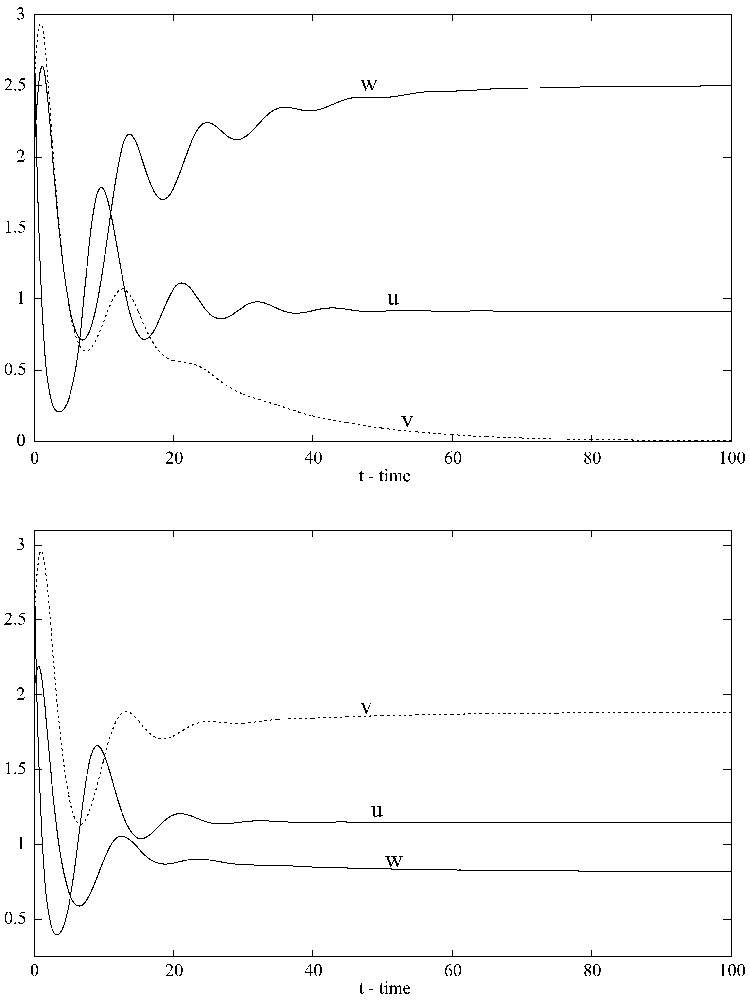
When G=0, the strong competitor with density w wins the competition and the weak competitor with density v tends toward extinction (top). When G=0.1, both competitors coexist and the solution converges to the positive equilibrium (1.1465,1.8837,0.8197) (bottom). XPPAUT was used for the simulations.
Introducing density-dependent mortality only for the competitor with density w (the stronger competitor) makes the system coexistent not only in the sense of uniform persistence, but also in the sense of existence of a globally stable positive equilibrium (Fig. 2). With the above parameter values and
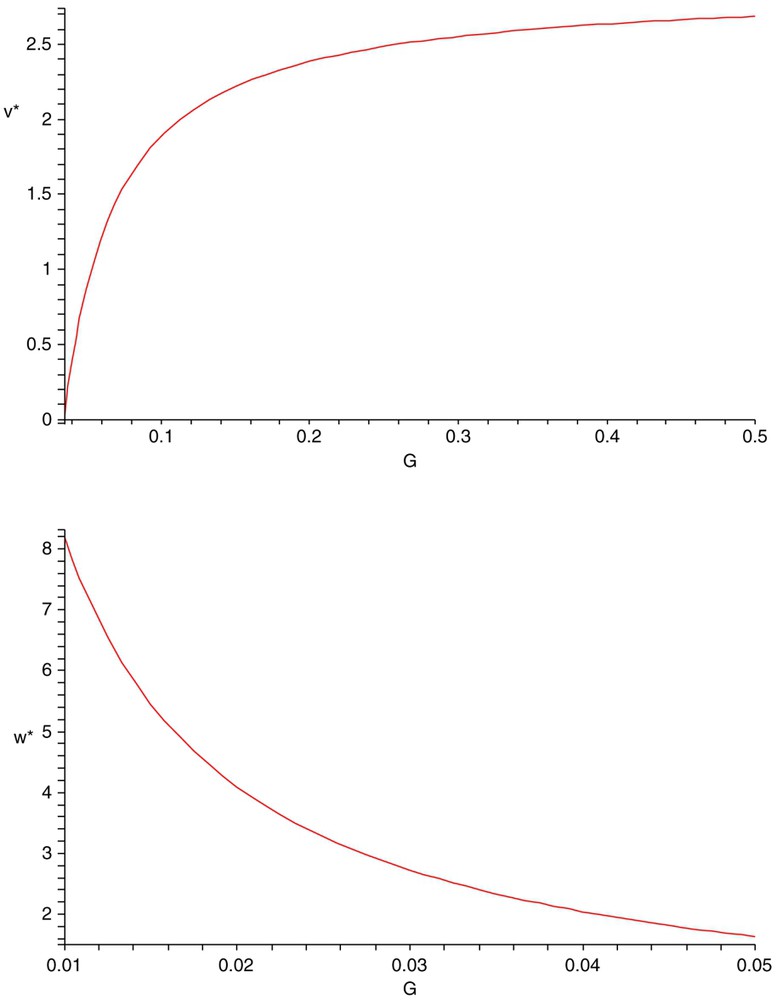
The steady-state value of the weak competitor with density v is an increasing function of G (top) and the steady-state value of the strong competitor with density w is a decreasing function of G (bottom). Here r=1.5, K=3, a=0.45, b=0.35, d=0.45, e=0.55, A=0.55, B=0.35, D=0.45, E=0.65.
Is the situation represented by model (1.4) a feasible one ecologically? It is a variation on the model of Tilman [44] for two consumers competing exploitatively for a single prey. Tilman's model predicts that the consumer that reduces the prey to the lower steady-state value will displace the other competitor. Model (1.4) includes the mechanism of density-dependent mortality, or biomass loss,
It is interesting to observe that, when the carrying capacity K of the prey is increased, the positive equilibrium loses its stability and a Hopf bifurcation occurs when K passes a critical value. With parameters given above, a supercritical Hopf bifurcation occurs when
It is also interesting to notice that, for the subsystem without the competitor v, that is, the predator-prey system with density-dependent mortality:
| (3.1) |
Acknowledgements
The authors are very grateful to the anonymous referee for his careful reading, detailed comments, and helpful suggestions. The first author would like to thank Prof. G. Bard Ermentrout and Dr. Rongsong Liu for their help in using XPPAUT for the numerical simulations.
Vous devez vous connecter pour continuer.
S'authentifier